
8.アポトーシス細胞の貪食と自己免疫の関わり
病気などで炎症が起こると組織細胞が傷害されたりしますが、それら細胞はネクローシスまたはアポトーシスによって死滅し、新たな細胞が出現することで正常組織の恒常性が保たれています。ネクローシスは主に、細胞内外の環境の悪化によって起こる細胞死であると考えられ、細胞膜が破れて、ネクローシスを起こした細胞中の消化酵素やサイトカインなどが流出し、炎症発生因子となって周囲細胞に重篤な影響を与え、ひどい場合はその組織が壊死することになります。一方、アポトーシスで死滅した細胞は、マクロファージ等の貪食細胞によって取り込まれて、消化されると考えられています。
長田チームはこれまでにアポトーシスの仕組みを次々と明らかにしてきました。最近、アポトーシス細胞において細胞膜内のリン脂質フォスファチジルセリン (PS)が細胞外に暴露することがマクロファージによる貪食の引き金になることを明らかにすると共に、PSを細胞外に暴露させる酵素(リン脂質スクランブラーゼ)の同定にも成功しました。さらに、アポトーシス細胞がうまく除去されなくなると自己免疫疾患が発症することを、PSに結合し死細胞の貪食を促進させる分子を欠損したマウスを用いて明らかにし、自己免疫疾患とアポトーシス細胞除去との関わりを指摘しています。また、血小板に発現するスクランブラーゼ(TMEM16F)遺伝子の変異が血友病の一種であるスコット症候群(Scott Syndrome)の原因であることを突き止めています。
【長田チーム】「リン脂質スクランブラーゼTMEM16FとXkr8の同定」Nature 2010, Science 2013
<ポイント>
- 細胞膜は外側と内側で成分が異なっていて、細胞内のみに存在するリン脂質フォスファチジルセリン (PS)の細胞外への暴露は種々の生体反応に重要である。
- PS暴露に関与するスクランブラーゼTMEM16F(血小板)とXkr8(アポトーシス細胞)を同定
- ある種の血友病やがん、自己免疫疾患などの疾患理解に貢献
動物細胞の細胞膜は二重膜を形成していますが、外側と内側では同一ではありません。リン脂質フォスファチジルセリン (PS) やフォスファチジルエタノールアミン (PE) は膜の内側にのみ存在するのに対し、フォスファチジルコリン(PC)は比較的外膜に多く存在します。しかし、この膜の非対称性は種々の局面で破綻します。例えば、細胞がアポトーシス注1) (細胞死)に陥ると死細胞の表面にはPSが暴露され、これをマクロファージが“eat me”シグナルと認識し、死細胞を貪食します。
一方、止血反応において出血した血管内皮に結合した血小板は、破壊された内皮細胞から放出された分子により活性化され、PSを暴露します。そして、このPSを介して凝固因子(Factor VIII, Factor V, Factor Xなど)が活性化され、血液凝固反応が進行します。このように、細胞膜におけるリン脂質の非対称性の崩壊は生体の種々の過程で重要な役割を果たしています。それにも関わらず、どのような機構で膜の非対称性が破綻するかその分子機構は殆ど解明されていません。
「血小板において止血の引き金となるリン脂質の暴露に関与する因子の同定-ヒトの遺伝病(スコット症候群)の原因遺伝子の同定」 Nature 2010
血小板が血液凝固をスタートさせるのに必要な膜蛋白質を同定しました。血液凝固が効率よくおこらないヒトの原因不明の病気 (Scott Syndrome、スコット症候群)の患者ではこの蛋白質を産生できなくなっていました。
リン脂質の非対称性を誘導する因子はスクランブラーゼ(Scramblase)と呼ばれ、カルシウム(Ca)によって活性化されると考えられていましたが、その実体は明らかでありませんでした。細胞内のCa濃度(100 nM) は細胞外(数mM)に比べて1万分の1以下です。京都大学 鈴木助教は膜にカルシウムを透過させる薬剤 (Ca-ionophore) でマウスB-リンパ球株(Ba/F3)を処理すると、PSが細胞膜上に暴露されること、この際、培養液中にCaが存在すると細胞は急速に死滅するが、Caが存在しないと細胞は死滅しないことを見いだしました。これは細胞外にCa が存在すると高濃度のCaが細胞内に流入し、細胞を殺したと考えられました。一方、細胞外にCaが存在しないと、Ca-ionophoreは小胞体の膜に作用し、小胞体内に蓄積されていたCa (数100 μM )を細胞質に流入させ、スクランブラーゼを活性化させたと考えられます(図1)。このような条件下で暴露されたPSはCa-ionophoreを除くと時間とともに減少しました。
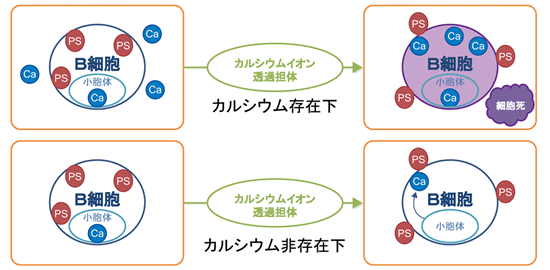
図1. Ca-ionophore(カルシウムイオン透過担体)処理により、Caが細胞質中に流入することでPSが暴露される。
(下)培養液中にカルシウムが存在していない状態でB細胞にCa-ionophore処理を行うと、小胞体内に蓄積された少量のカルシウムが細胞質中に流入し、スクランブラーゼの活性化によりPSが細胞外に暴露されるが、Ca-ionophoreを除くと暴露されたPS量が減った。
鈴木助教は、細胞をCa非存在下でCa-ionophoreで処理するとPSが一過的に暴露される特徴を用いて、PSをよく暴露する細胞を樹立しました。すなわち、Ca非存在下でBa/F3をCa-ionophoreで処理、PSを最もよく暴露した細胞 (全体の1?2%) をFACS(Fluorescence-activated cell sorting) 法により分別、収集しました。この細胞を培養、増殖させ、再びCa-ionophoreで処理、1-2%の分画を採集します。この操作を19回繰り返しました。その結果、親株であるBa/F3細胞より100倍以上強くPSを暴露する細胞株が樹立されました。次いでこの細胞株から動物発現ベクターを用いてcDNAライブラリーを構築、 これをBa/F3細胞株に導入しPSの暴露に関与する遺伝子を同定しました。
単離されたcDNAは、TMEM16Fと呼ばれる8個の膜貫通領域を持つ膜蛋白質に1個のアミノ酸置換が導入された変異蛋白質をコードしていました (図2)。このcDNAを発現させたBa/F3細胞は構成的にPSを暴露しました。一方、野生型のTMEM16FをBa/F3細胞に発現させ、Caで刺激するとその細胞は親株に比べ急速にPSを細胞外に暴露しました。また、shRNA(small hairpin RNA) 法によりBa/F3細胞で発現されているTMEM16F mRNA量を減少させると、Caに応答したPSの暴露は顕著に抑制されました。Scramblaseは、PSばかりでなく、他のリン脂質の外膜、内膜の移動も促進するとされています。実際、変異型TMEM16Fを発現するBa/F3細胞は本来内側にあるべきPEを構成的に細胞外に暴露し、また通常は外膜に大部分存在するPCを内膜へと移動させました。また野生型のTMEM16Fを発現する細胞ではこれらリン脂質の移動が顕著に促進され、shRNAによりTMEM16Fの発現が減少した細胞ではリン脂質の移動が抑制されました。以上より、TMEM16FはCaに依存してリン脂質を内膜、外膜の間でスクランブルさせる因子、スクランブラーゼと結論しました。
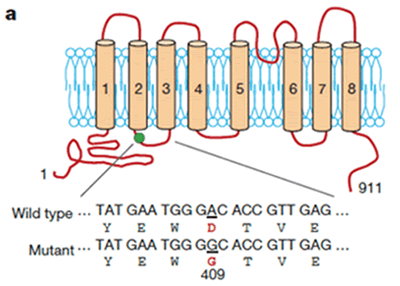
図2 Scramblase TMEM16Fの構造
さて、第X血液凝固因子(スチュワートブラウワー因子)やトロンビンが活性化されず、血液凝固が効率よく進まないスコット症候群(Scott Syndrome)と呼ばれる病気が知られています。この患者さんの血小板や赤血球、B-リンパ球をCaで刺激してもPSが細胞外に暴露されないことからスクランブラーゼに欠損があると考えられました。1994年、アメリカ・ウイスコンシン Sims 博士の研究室に留学していた小島寛博士(現 茨城県立中央病院・腫瘍内科)はこの患者さん、および患者さんの両親から調製したリンパ球をウイルスを用いて不死化し、細胞株を樹立しました。その細胞株がPeter J. Sims博士(現在Rochester University Medical Schoolの教授)の研究室に保管されていたことから、これらの細胞を京都大学に送っていただき、鈴木助教はこれら細胞におけるTMEM16F 遺伝子、TMEM16F mRNAを解析しました。TMEM16F遺伝子は20個のエクソンから成り立っています。患者さんのTMEM16F mRNAでは13番目のエクソンが欠質、そのためこの蛋白質は分子のほぼ半ばで途切れていました (図3)。mRNAの前駆体からmRNA が産生される際、残すべきエクソンは“AG----GT”の配列で囲まれています。スコット症候群の患者さんのTMEM16F 遺伝子ではエクソン13の5’側においてAGであるべき配列がATに変異していました。そのため、エクソン13はスキップされ、エクソン12はエクソン14に直接、結合しました。その結果、mRNA上での蛋白合成のフレームがずれ、蛋白質への翻訳はエクソン14上で終結、ほぼ半分の大きさの蛋白質しかできなくなっていました。患者さんの両親はこの変異を持つ遺伝子と正常な遺伝子を1個ずつもちますが、患者さんは2個の遺伝子とも変異型であり、このためスクランブラーゼが欠損、血小板はPSを暴露できないと考えられます。
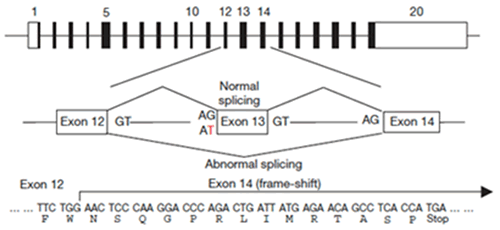
図3. スコット 症候群の患者に見いだされたTMEM16F遺伝子の変異
私達は血小板が血液凝固を開始する際のシグナル、PSの細胞表面への暴露を引き起こす因子 (TMEM16F) を同定しました。そして、スコット症候群の一人の患者さんにおいて、この遺伝子に変異があることを見いだしました。今後はスコット症候群の他の患者さんもTMEM16F遺伝子に変異が存在するか、血液凝固に欠陥を持つ患者さんでこの因子の変異で説明できる患者さんが存在するか検討する必要があるでしょう。また、今後、TMEM16Fの活性を促進あるいは抑制する薬剤が開発できれば、これらは血液凝固の促進、あるいは血栓形成阻止の薬として使われる可能性もあるでしょう。最後に、細胞膜の非対称性の破綻、PSの暴露は死細胞の貪食、筋肉細胞や骨芽細胞の成熟過程にも関与しています。今回の成果は今後これらの過程を理解する上でも役立つでしょう。
<共同研究者>
鈴木淳 京都大学大学院医学研究科 分子生体統御学 医化学分野 助教
梅田真郷 京都大学大学院工学研究科 合成・生物化学専攻 教授
Dr. Peter J. Sims, Professor, Dept. of Pathology & Laboratory Medicine, University of Rochester Medical Center (ロチェスター大学医学部 病理学教室教授)
長田重一 京都大学大学院医学研究科 分子生体統御学 医化学分野 教授
<原論文情報>
Suzuki J, Umeda M, Sims PJ, Nagata S. Calcium-dependent phospholipid scrambling by TMEM16F. Nature 2010
「アポトーシス時のリン脂質暴露に関与する因子の同定」Science 2013
アポトーシス(細胞死)を起こした細胞はマクロファージなどの貪食細胞に貪食されます。この際、死細胞は「eat me」シグナルとしてフォスファチジルセリン(PS)をその表面に暴露します。鈴木淳 医学研究科助教、長田重一 同教授、今西英一 同教務補佐員らのグループは、H. Robert Horvitz アメリカマサチューセッツ工科大学教授、Daniel P. Denning 同博士研究員のグループと共同で、このフォスファチジルセリン暴露の過程に関与している膜蛋白質を同定しました。
2010年、鈴木助教はフォスファチジルセリンを強く暴露するマウス細胞株(Ba/F3-PS19)を樹立し、この細胞株からCa2+に応答してPSを暴露させる蛋白質(TMEM16F)を同定しました。そして、血友病の一種、Scott Syndromeと呼ばれる患者はこの遺伝子に変異を持つことを示しました。実際、TMEM16F遺伝子を欠損したマウス細胞ではCa2+に依存したリン脂質のスクランブリングは全く起こりませんでした。一方、アポトーシス時のリン脂質のスクランブリング、PSの暴露はTMEM16F欠損細胞でも、野生型の細胞と同じ効率で進行しました。このことから、アポトーシス時にリン脂質のスクランブリングを引き起こす分子は別に存在すると考えられました(鈴木助教ら J. Biol.Chem.288, 13305, 2013)。
そこで、鈴木助教は上記Ba/F3-PS19細胞株からのcDNAライブラリーを再度スクリーニング、PSを暴露させる能力を持つcDNAを単離しました。単離されたcDNAはXkr8と呼ばれる6個の膜貫通領域を持つ膜蛋白質をコードしていました(図4)。マウスT-リンパ球株WR19LにこのcDNAを発現させると、Fasリガンドによるアポトーシス時のPSの暴露が顕著に増強されました。一方、Xkr8遺伝子をノックアウトしたマウスから樹立した繊維芽細胞ではアポトーシス時のPSの暴露が完全に失われました。スクランブラーゼは、PSばかりでなく、他のリン脂質の外膜、内膜の移動も促進するとされています。実際、Xkr8を発現する細胞にアポトーシスを起こすと、PSばかりでなく、フォスファチジルエタノールアミンも細胞外に暴露され、また通常は外膜に存在するフォスファチジルコリンが内膜へと移動しました。ところで、Xkr8は線虫のCED8と呼ばれる遺伝子とそのアミノ酸配列が20%相同です。Horvitz教授らは、CED8を欠損した線虫ではアポトーシスを起こした細胞の表面にPSが暴露されないことを確認しました。以上より、Xkr8/CED8はアポトーシス時、リン脂質を内膜、外膜の間でスクランブリングさせる過程に関与していると結論づけました。
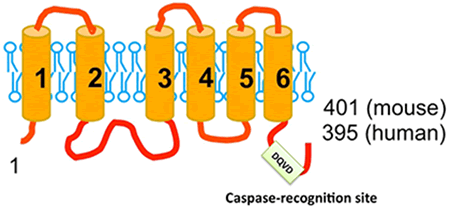
図4:Xkr8の構造とカスパーゼ認識配列
それでは、Xkr8はアポトーシス時にどのように活性化されるのでしょうか。アポトーシスはカスパーゼと呼ばれる蛋白質分解酵素によって実行されますが、PSの暴露もカスパーゼを阻害すると起こりません。そこで、Xkr8蛋白質のアミノ酸配列にカスパーゼの認識配列が存在しないかを調べると、ヒトやマウスのXkr8蛋白質のC-末端近傍にカスパーゼ3によって切断される配列が認められました(図4)。そして、実際、アポトーシス時にこの蛋白質が切断されること、切断されない変異体ではリン脂質をスクランブリングできないことを見いだしました。すなわち、Xkr8は健康な細胞内では活性のない前駆体として存在し、アポトーシスが誘導されるとカスパーゼにより切断、活性のある蛋白質に変換されると結論づけました。
アポトーシス時のPSの暴露はほとんどの細胞で起こりますが、ある種のヒト白血病細胞(RajiやPLB985細胞)ではPSは暴露されず、マクロファージがその死細胞を貪食することもありません。そこで、これら白血病細胞でのXkr8遺伝子を調べたところ、その遺伝子構造は正常であるにも関わらず、ほとんど発現されていませんでした。一方、Xkr8遺伝子のプロモーターはCGに富んでおり(CpG islands)、RajiやPLB985白血病細胞では健常人の白血球細胞と異なり、そのシトシン塩基が高い頻度でメチル化されていました(図5)。このことから、これらヒト白血病細胞ではXkr8遺伝子のプロモーターのCpG残基がメチル化され、これによってその発現が抑制されていると結論しました。実際、PLB985細胞を脱メチル化試薬DAC(5'-aza-2'-deoxycytidine)で処理するとプロモーター領域のメチル基は除去され、Xkr8遺伝子が発現、アポトーシス時のPSの暴露も観察されました。
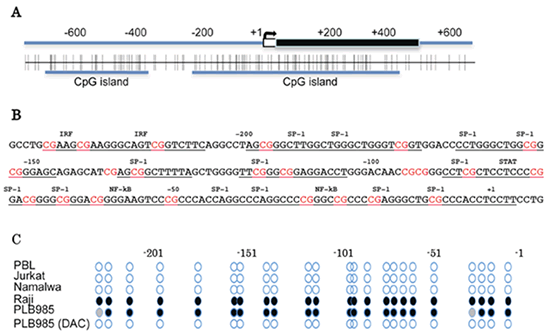
図5:ヒトXkr8遺伝子プロモーター領域のCpG islandsと白血病細胞でのメチル化
今回本研究グループは、アポトーシス細胞において「eat me」シグナルとして作用するフォスファチジルセリンの暴露に関与する膜蛋白質(Xkr8)を同定しました。この蛋白質は増殖している細胞中では活性のない前駆体として存在し、アポトーシスがおこるとカスパーゼによって直接、切断活性化されます。フォスファチジルセリンは貪食細胞が死細胞を貪食するためのシグナルであり、フォスファチジルセリンが提示されないと、死細胞は貪食されず、細胞膜が破裂、いわゆるネクローシス状態に陥ると考えられます。ネクローシスに陥った細胞からは核やDNA、ミトコンドリアなどが放出され、自己抗体の産生を介して、SLE(Systemic Lupus Erythematosus、全身性エリテマトーデス)様の自己免疫疾患を発症させる可能性があると思われます。SLEなどのヒトの自己免疫疾患がXkr8遺伝子の作用不全で起こっているかどうかを調べる必要があります。ところで、ある種のがん細胞において、がん抑制遺伝子の発現がその遺伝子プローモーターに存在するCpG islandsのメチル化によって抑制されていることが知られています。今回、本研究グループは2種のヒト白血病細胞でXkr8遺伝子のプロモーターがメチル化され、その発現が抑制されていることを見いだしました。このようながん細胞はアポトーシスに陥っても貪食されず、ネクローシスに陥った細胞から放出された細胞内分子により、強い炎症反応が惹起される可能性があります。このことががんの進行を引き起こす可能性がないかどうかが今後の課題です。最後に、Ca2+に応答してリン脂質をスクランブルするTMEM16Fとアポトーシス時に作用するXkr8はアミノ酸配列は全く異なっています。これらの分子がどのようにリン脂質をスクランブルさせるのか、その分子機構は大変興味深いものです。
<用語解説>
- 注1) アポトーシス
- 細胞の死に方の1つの形態。プログラムされた細胞死ともいわれ、個体にとって余分な細胞や、危険な細胞が個体の持つ制御機構に従って自殺する時の細胞の死に方。生理的にも毎日、全身の多数の細胞がアポトーシスにより死んで新しい細胞によって置き換わっている。細菌などの感染による炎症反応によってもアポトーシスによる細胞死が起こる。 これに対し、栄養不足などにより細胞質が変化することによって、細胞が死に至る場合はネクローシス(壊死)という。
<共同研究者>
本研究は京都大学大学院医学研究科 鈴木淳 助教、長田重一 教授、今西英一 教務補佐員らのグループが、H.Robert Horvitz アメリカマサチューセッツ工科大学教授、Daniel P.Denning 同博士研究員のグループと共同で行いました。
<原論文情報>
Suzuki Jun, Denning Daniel P., Imanishi Eiichi, Horvitz H. Robert, Nagata Shigekazu.
Xk-Related Protein 8 and CED-8 Promote Phosphatidylserine Exposure in Apoptotic Cells. Science, 2013