
- 【吉村チーム】「免疫反応を抑える細胞が作られる新たな仕組みを発見 」Nature Immunology 2013
- 【竹田チーム】「マウスの腸内で自然免疫細胞が炎症を抑える新たな仕組みを解明」PNAS 2012
4.T細胞活性制御のしくみ
免疫反応は有害な病原体を排除するために重要ですが、炎症反応を伴うので適切な時期に終息しないと、自分自身の体を攻撃することになります。そのために、免疫系には免疫応答を抑制制御する様々なしくみが備わっています。例えば、免疫応答が終わる際には大部分の抗原特異的免疫細胞はアポトーシスと呼ばれるプログラムされた細胞死が誘導されたり、制御性T細胞と呼ばれる免疫反応を抑制する免疫細胞が働いたりします。制御性T細胞は、また、自分自身の蛋白質(自己抗原)を認識する免疫細胞を不活性な状態(免疫寛容)のままに維持する重要な役割も果たします。
吉村チームは、制御性T細胞の誘導に関わる新たな核内受容体を同定し、免疫疾患の制御法の開発を目指しています。また、竹田チームは、自然免疫系に関わる細胞の中に、T細胞の活性化を抑制する新たな細胞、制御性ミエロイド細胞(Mreg)の存在を見出しています。
【吉村チーム】「免疫反応を抑える細胞が作られる新たな仕組みを発見 」Nature Immunology 2013
<ポイント>
- 過剰な免疫反応を抑制する制御性T細胞(Treg)注1)が作られる仕組みは不明だった。
- 核内受容体Nr4a注2)を適度に活性化すると、Tregが作られることを発見。
- 関節リウマチなどの自己免疫疾患注3)や花粉症、ぜんそくなどのアレルギー性疾患の新しい治療法に期待。
免疫システムは我々の身体を多種多様な病原体から守っています。病原体は、細菌・ウイルス・寄生虫・真菌、さらにその中にも数限りない種類が存在します。それら異なった種類の病原体を排除するために、生体は炎症などを伴う適切な免疫反応を作動させますが、その司令塔としての役割を果たしているのがヘルパーT細胞(CD4陽性T細胞注4))です。ヘルパーT細胞は胸腺で生まれ、胸腺の外(末梢)に出て身体を循環し、身体に侵入した病原体の種類に応じてTh1、Th2、Th17の3種類のいずれかの炎症を引き起こすエフェクターT細胞注5)に分化し、それぞれの病原体の排除に最適な免疫反応を誘導します(図1)。一方、免疫反応は適切な時期に終息しなければなりません。また、自己のたんぱく質や食物には反応してはいけません。もし異常に免疫系が活性化され続けたり、自己のたんぱく質や食物や花粉などの無害な物質と反応すると、関節リウマチや炎症性腸疾患などの自己免疫疾患や花粉症などのアレルギーを引き起こします。そこで、ヘルパーT細胞のなかにはエフェクターT細胞を抑制し免疫反応を適切に制御するT細胞、制御性T細胞(Treg)が存在します。Tregはさまざまなメカニズムで免疫反応を抑制し、生体の恒常性(ホメオスターシス)の維持に重要な役割をはたしています。Tregの異常はエフェクターT細胞の過剰な活性化を招き、自己免疫疾患やアレルギー性疾患を引き起こします。逆にTregを人為的に増強できればそれら疾患の治療に結びつくと考えられます。また臓器移植の拒絶反応にもエフェクターT細胞が寄与しており、Tregの増強は臓器移植の拒絶反応の抑制にも効果的であると考えられています。しかしTregそのものがどのように発生するのかは長らく不明で、Treg発生のメカニズムの解明は現代免疫学における大きな課題の1つでした。

図1 ヘルパー(CD4陽性)T細胞の分化とTregによるエフェクターT細胞の抑制
ヘルパーT細胞は、胸腺で前駆細胞から発生・分化を遂げますが、この過程で抗原を感知するT細胞受容体(TCR)注6)がランダムな遺伝子組み換えによって数千万種類も形成され、外からのさまざまな病原体の排除に備えます。T細胞を増やしたり生存させたりするためには、外からの抗原がない状態でもTCRが適度の刺激を受ける必要があります。このため胸腺では、抗原提示細胞注7)に提示された自己抗原(主に自己細胞のたんぱく質)がTCRを弱く刺激します。この結果おのおののT細胞が異なったTCRを持って、さまざまな抗原の排除に対応できる免疫システムを構築できるようになります。しかし、ランダムな遺伝子組み換えによってさまざまなTCRを形成するので、当然自分自身の細胞が持つたんぱく質を抗原と認識して強く反応するTCRを持つT細胞が発生する可能性があります。胸腺での発生過程では、これらの自己抗原に対して強い反応を起こすT細胞は強いTCR刺激が入り、アポトーシス(細胞死)注8)と呼ばれる機構によって死亡し除去されます。この現象をネガティブセレクションと呼びます(図2)。さらに、それよりは少し弱いが自己抗原をある程度強く認識するTCRを持つT細胞が死なずにTregになるのだろうと考えられています(図2、Treg)。それよりもさらに弱く自己抗原を認識するT細胞が、病原体排除に機能する一般的なT細胞(ナイーブT細胞注9))として胸腺の外に出て末梢を循環します(図2、ナイーブT細胞)。この一般的なT細胞が末梢で抗原に出会うと先に述べたエフェクターT細胞になります。また、自己抗原を全く認識できない細胞はアポトーシスが誘導され死滅(図2、細胞死)します。このように、胸腺においてヘルパーT細胞は、主に自己抗原に対する親和性(反応性)によりその運命を決定づけられます。この仕組みが免疫システムの暴走を防ぎ、恒常性を維持する基盤となります。しかし、Tregを産み出す分子メカニズムは多くが未解明で、人為的にコントロールすることも不可能でした。
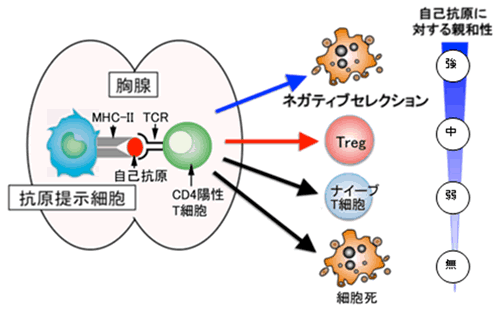
図2 胸腺におけるヘルパー(CD4陽性)T細胞発生の分化
吉村チームは、これまでの研究で核内受容体ファミリーに属するNr4a2というたんぱく質がTregの性質を決める重要な転写因子であるFoxp3注10)の発現を誘導する能力があることを見いだし、報告を行いました(図3)(参考文献)。さらに、Nr4a2やその仲間はTCRの活性化によって多く生産される分子であることから、Nr4a2が胸腺でのT細胞の運命決定、特にTregを生み出す鍵となる分子であると考え、その役割の解明を進めることにしました。
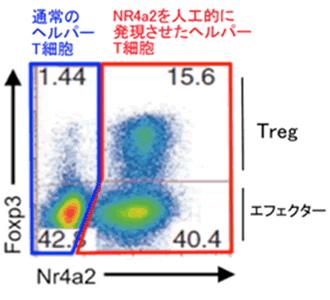
図3 Nr4a2強制発現によるTreg主要転写因子Foxp3の誘導
本研究グループはNr4a2の役割を解明するために、まずマウスのNr4a2のみを欠損させたところ、免疫反応が促進されるものの、胸腺でTregは正常に発生し、さらにそのマウスは自己免疫疾患を発症しませんでした。そこで、Nr4a2には、Nr4a1、Nr4a3という良く似た仲間(ファミリー分子)が存在することに着目し、この仲間が互いに同様の機能を持ってその機能を補い合っていると考え、Nr4a1、Nr4a2、Nr4a3全てをT細胞から欠損させたマウス(Nr4a-TKOマウス:Nr4a triple knockout mouse)を作製し、解析を行いました。
解析の結果、予想通り、Nr4a-TKOマウスでは胸腺・末梢ともにTregがほぼ完全に存在しないことが明らかとなりました(図4A)。その結果、Nr4a-TKOマウスは激しい全身性の自己免疫疾患を発症し、生後3週間以内に死亡することが明らかとなりました(図4B)。従って、Nr4aファミリー分子がTregの発生に必須であることが証明されました。またNr4a-TKOマウスの症状は、過去の文献との比較により、Tregのみを欠損しているマウスよりも重篤であり、さまざまな解析の結果からもTregの分化異常に加えて、ネガティブセレクションの異常が関与している可能性が強く考えられました。さらに解析を重ねたところ、Nr4a1、Nr4a2、Nr4a3は同様の機能を持ち、互いに補い合ってTregの分化とネガティブセレクションに機能していることが明らかとなりました。
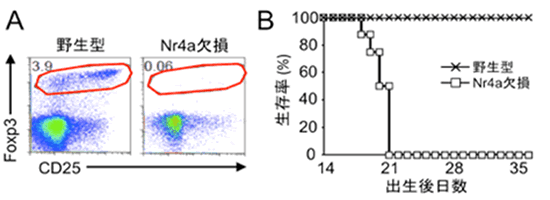
図4 Nr4a欠損マウスの解析
(B)Nr4a欠損マウスは全身性の自己免疫疾患により生後3週間以内に死亡する。
次に、Nr4aがヘルパーT細胞の運命決定を担う因子であると考え、その検証を試みました。実験にはOT-IIと呼ばれるTCRを持つT細胞を用いました。マウスの胸腺内にはOT-II TCRと強く結合する自己抗原が存在しないため、OT-II TCRを持つT細胞はTregに分化するために必要なTCRの刺激を受けることができず、OT-II TCRを持つTregは存在しないことが明らかとなっています。そこで、このOT-II TCRを持つヘルパーT細胞に、遺伝子操作によって任意の強さで人工的に活性化させたNr4aを作用させ、ヘルパーT細胞の運命がどのように決定されるかを解析しました。その結果Nr4aを強力に活性化させるとOT-II TCRを持つヘルパーT細胞にアポトーシス(細胞死・ネガティブセレクション)が誘導され、中程度の活性化を起こすとTregが誘導されることを見いだしました(図5)。このことは、TCRと自己抗原との親和性が強い時は、Nr4aも強く活性化され、アポトーシス誘導遺伝子の発現が誘導されてネガティブセレクションへ、TCRと自己抗原との親和性が中程度の時は、Foxp3の発現が誘導されTregへとヘルパーT細胞の運命が決定づけられていることが分かりました(図6)。
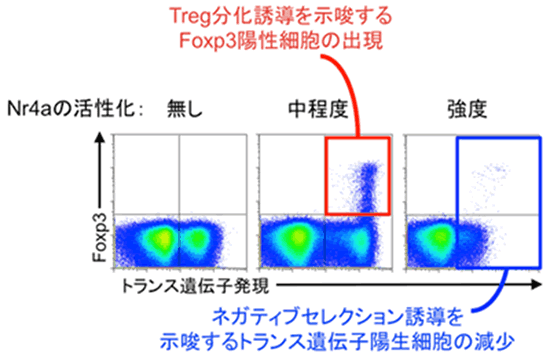
図5 Nr4aはCD4陽性T細胞の自己抗原に対する親和性と運命決定を結ぶキーファクターとして機能する。
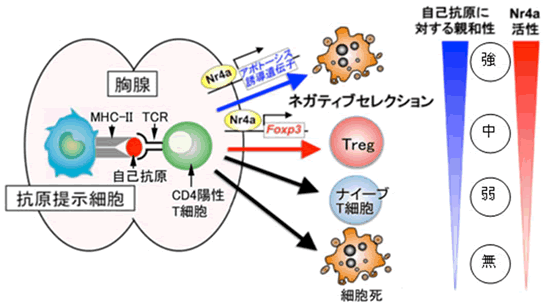
図6 胸腺ヘルパーT細胞運命決定におけるNr4a機能のモデル図
以上の研究により、胸腺におけるヘルパーT細胞の発生において、Nr4aがヘルパーT細胞と自己抗原との親和性とヘルパー細胞の運命決定を結ぶ鍵となるたんぱく質であることを証明しました。また、Nr4aを強度に活性化するとネガティブセレクションが、Nr4aを中程度に活性化することによって普通はTregにならないT細胞から人為的にTregが作られることを示しました。
本研究によって、Nr4aはヘルパーT細胞の運命決定を担う鍵となり、Tregを作る重要な役割を果たす因子であり、自己免疫疾患の抑制において中心的な役割を担っていることが明らかとなりました。このたんぱく質の活性化によって、免疫反応を抑制するTreg細胞を人為的に誘導することも可能になると考えられます。特にNr4aは核内受容体と呼ばれる分子の性質を持っており、活性化させる低分子化合物(リガンド)の存在も予想されます。今後さらなる研究からNr4aの活性化による自己免疫疾患、アレルギー疾患などの新規治療法の樹立が可能になると期待されます。また臓器移植の際の拒絶反応抑制法の開発も期待できます。さらに、ヒトの炎症性疾患とNr4a遺伝子異常の関連性も明らかにされることで、免疫難病の発症の仕組みが解明されることが期待されます。
<用語解説>
- 注1) 制御性T細胞(Treg:ティーレグ)
- CD4陽性ヘルパーT細胞の一種で、免疫抑制機能を持つ。自己に対する免疫応答を抑制するほか、IL-10やTGFβなどの抗炎症性サイトカインを分泌し、過剰な炎症を抑制する機能も持つ。主に胸腺で作られ、免疫システムの負の制御において中心的な役割を担っており、その分化や機能発現の異常は自己免疫疾患やアレルギーをはじめさまざまな免疫疾患の引き金となる。
- 注2) 核内受容体
- DNAの転写を調節するたんぱく質、いわゆる転写因子の一種であるが、ホルモンなどのリガンドにより活性調節を受けるドメインを分子内に持つ。ヒトでは48種類の核内受容体が存在し、スーパーファミリーを形成する。エストロゲン受容体やビタミンD受容体もこれに属し、それぞれ女性ホルモンとビタミンDにより活性化を受ける。Nr4aもこのファミリーに属する。
- 注3) 関節リウマチや炎症性腸疾患などの自己免疫疾患
- 自己免疫疾患とは、免疫系が自分自身の正常な細胞や組織に対して過剰に反応し、攻撃を加えることにより発症する疾患の総称である。関節リウマチでは手足の関節、炎症性腸疾患では腸管壁で過剰な免疫反応が起こり、炎症が生じる。国内患者数は70万人と言われている。炎症性腸疾患の患者数は著明な増加傾向にあり、国内患者数は潰瘍性大腸炎とクローン病をあわせて15万人を超している。
- 注4) CD4陽性T細胞
- CD4蛋白分子を細胞表面に発現するT細胞。細胞障害性T細胞の機能発現の誘導、B細胞の分化成熟や抗体産生の誘導、貪食細胞の活性化を誘導し免疫応答を正に制御するエフェクターT細胞と、免疫応答を負に制御する制御性T細胞に大別される。
- 注5) エフェクターT細胞
- 免疫系を活性化するT細胞。特にヘルパーT細胞では主にTh1、Th2、Th17の3種類が知られている。それぞれインターフェロンγ(IFNγ)やインターロイキン17(IL-17)など特徴的なサイトカインを放出し、感染防御や免疫応答に関わるほか、自己免疫疾患、アレルギー、がんでも重要な働きをする。
- 注6) T細胞受容体(TCR:T cell receptor)
- T細胞表面に発現している分子であり、MHC-ペプチド抗原複合体を認識する。ヘルパーT細胞が発現するTCRはMHC-IIに掲示された抗原を認識する。末梢において、T細胞受容体は下流にシグナルを伝え、T細胞の機能発現や増殖が誘導され、エフェクターへと成熟させる。胸腺においてはT細胞受容体からのシグナル伝達は、T細胞発生分化の運命決定を担う。
- 注7) 抗原提示細胞
- 樹状細胞、マクロファージ、B細胞、胸腺上皮細胞など。外来性抗原をヘルパーT細胞に提示する機能を持つ蛋白分子を主要組織適合性抗原クラスII(MHC-II:major histocompatibility antigen class II)と呼び、抗原提示細胞の細胞表面にあって、抗原と結合してTCRを刺激する。胸腺では自己蛋白質の一部を提示し、末梢においては主に病原体などの外来の抗原を提示する。 また、キラーT細胞にウイルス抗原などの内在性抗原を提示する蛋白分子をMHC-IIと呼ぶ。
- 注8) アポトーシス(細胞死)
- 外傷性ストレスで起こる細胞死でなく、細胞内に備わる分子機構により誘導される、管理・調節された細胞死。
- 注9) ナイーブT細胞
- T細胞は胸腺においては自己抗原を弱く認識して生存シグナルを受け発達し胸腺の外に出た段階でナイーブT細胞と呼ばれる。胸腺の外に出るとナイーブCD4陽性T細胞は病原体などの抗原を認識し、病原体の種類や周囲の環境に応じてエフェクターT細胞へと分化誘導され、最適の免疫応答を引き起こす。
- 注10) Foxp3(フォックス ピー スリー)
- Tregの分化・機能発現・分化状態の維持全てにおいて必須の役割を担う転写因子。発現はTregにほぼ特異的であるため、Tregを同定する際のマーカー分子としても用いられる。免疫抑制機能を持つ蛋白質の発現を誘導し、一方で炎症に関わる蛋白質の発現を抑制する。
<原論文情報>
“Nr4a receptors are essential for thymic regulatory T cell development and immune homeostasis”(Nr4a受容体は胸腺におけるTregの発生と免疫恒常性の維持に必須である)
Nat Immunol. 2013 Mar;14(3):230-7. doi: 10.1038/ni.2520. Epub 2013 Jan 20.
Sekiya T, Kashiwagi I, Yoshida R, Fukaya T, Morita R, Kimura A, Ichinose H, Metzger D, Chambon P, Yoshimura A.
<参考文献>
Sekiya, T. et al. “The nuclear orphan receptor Nr4a2 induces Foxp3 and regulates differentiation of CD4+ T cells.”, Nat Commun 2011;2:269. doi: 10.1038/ncomms1272.
Sekiya T Kashiwagi I, Inoue N, Morita R, Hori S, Waldmann H, Rudensky AY, Ichinose H, Metzger D, Chambon P, Yoshimura A.
【竹田チーム】「マウスの腸内で自然免疫細胞が炎症を抑える新たな仕組みを解明」PNAS 2012
<ポイント>
- 日本で急激な増加傾向にある炎症性腸疾患(IBD)研究は炎症性T細胞を中心としていたが未解明な部分が多い
- 炎症性T細胞を直接抑制させる制御性自然免疫細胞(Mreg細胞)を同定
- IBDをはじめとする多くの自己免疫疾患の治療法開発に期待
免疫細胞注1)は、細菌やウイルスといった異物を認識して攻撃・排除することによって、生体を防御する仕組みにおいて重要な役割を担っています。自然免疫細胞注2)は体内に侵入してきた病原体を消化して、その構造の特徴を獲得免疫細胞注3)であるT細胞に伝えることで獲得免疫系を活性化します。しかし、腸管内には100兆個の腸内常在細菌や食べ物由来の抗原といった異物が常時存在するため、腸管の粘膜において免疫細胞が過剰に活性化すると、腸管の組織を破壊し、腸炎の発症につながる恐れがあります。そこで、腸管粘膜に存在する免疫細胞は腸内の異物に対して普段は反応しない免疫寛容注4)の状態である必要があります。
腸管組織における免疫寛容の破綻は、日本で急激な増加傾向にある炎症性腸疾患注5)(IBD)の原因となることが報告されています。IBDの病因および発症機序は未解明な部分が多く、病態を根本的に改善する方法は確立されていません。IBDの特徴として炎症を導くT細胞が異常に増加していることが挙げられるため、今までのIBDの病因や発症機序の解明への取り組みはT細胞が主体である獲得免疫系を中心としたものでした。しかし、近年、自然免疫細胞に異常があると、獲得免疫系の活性化を抑制する仕組みが破綻することが明らかとなり、腸組織の恒常性維持における自然免疫細胞の役割に注目が集まっています。
腸内での免疫寛容状態の維持や、腸炎の抑制に重要な役割を果たす獲得免疫細胞としては、制御性T細胞(Treg(ティーレグ)細胞)が知られています。Treg細胞は、炎症を誘導するT細胞(炎症性T細胞)の活性化を抑制しています。これまでに、腸管粘膜に存在する自然免疫細胞の中にTreg細胞を誘導することによって、間接的に炎症を抑制しているものがいくつか報告されていました。しかし、炎症性T細胞を直接的に抑制する自然免疫細胞は報告されていませんでした。
そこで竹田チームは、Treg細胞を介さず直接的に腸炎を改善することができる自然免疫細胞が存在するのかを調べることにしました。
本研究グループは、T細胞の増殖を抑制する自然免疫細胞を探すために、マウスの大腸粘膜から回収した自然免疫細胞のうち最も大きな細胞集団に着目しました。この細胞を調べたところ、3つのグループがあることが確認され(図1A)、そのうちの1つの細胞が、T細胞の増殖を抑制することが分かりました(図1B)。さらに調べると、この細胞は細胞接着分子注6)を多く発現するほか、炎症性のT細胞に良く結合することも明らかになりました。
次に、この細胞の生体内での機能を解析するために、IBDモデルマウスの腹腔にこの細胞を投与しました。その結果、この細胞が腸管組織で炎症性T細胞の増殖を抑制して腸炎を抑えていることが明らかとなりました(図1C)。本研究グループは、この細胞を、Treg細胞を誘導するのではなく細胞上にある細胞接着分子によって、炎症性T細胞と直接結合して増殖を抑制することから、新規の自然免疫細胞集団として、「制御性ミエロイド細胞(regulatory myeloid cell;Mreg細胞)」と名付けました。
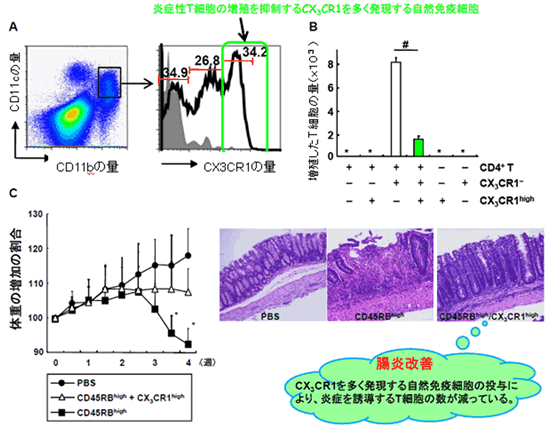
図1 大腸に存在するCX3CR1high CD11b+ CD11c+ 細胞は腸炎抑制能を示す
- A 野生型マウス(C57BL/6Jマウス)より回収した大腸粘膜固有層細胞をCD11c、CD11b、CX3CR1で染色すると、CD11b+ CD11c+ 細胞にはCX3CR1の発現量が異なる3つの集団が存在する。左図の黒枠内はCD11b+ CD11c+ 細胞を示す。
- B CX3CR1・ CD11b+ CD11c+ 細胞により誘導されるT細胞の増殖は、CX3CR1high CD11b+ CD11c+ 細胞を添加すると抑制される。
- C IBDモデルとして、SCIDマウスにCD4+ CD45high T細胞を3x105個移入。同時に、大腸のCX3CR1high CD11b+ CD11c+ 細胞を3x105個投与すると腸炎が抑制される。PBSは緩衝液で、コントロール実験として用いた。
Mreg細胞による腸炎の抑制に関与する分子機構をさらに明らかにするため、遺伝子発現解析を行った結果、Mreg細胞では腸管粘膜にたくさん存在する抗炎症性サイトカインのIL-10や、Stat3という転写因子注7)によって発現が誘導される遺伝子が多数存在していることが分かりました。そこで、Mreg細胞による炎症性T細胞の増殖抑制にはIL-10やStat3が必要であるという仮説を立てました。
この仮説を実証するため、IL-10またはStat3の機能を破壊したMreg細胞を炎症性T細胞と一緒に培養したところ、炎症性T細胞の増殖は抑制されませんでした。さらに、Stat3の機能を破壊したMreg細胞をIBDモデルマウスの腹腔に投与しても腸炎が抑制されないことから、Mreg細胞の炎症性T細胞の増殖抑制や腸炎抑制には、IL-10やStat3が必要であると考えられました(図2A)。さらに詳細に解析した結果、T細胞の活性化に必須のCD80やCD86というたんぱく質の発現が、Mreg細胞ではIL-10やStat3によって抑制されていることが明らかとなりました。IBDモデルマウスの腹腔に正常なMreg細胞を注射によって投与したところ、腸炎の症状が改善されました(図2B)。これらの結果により、Mreg細胞はIL-10やStat3に依存して腸炎の発症抑制に重要な役割を果たすことが分かりました。
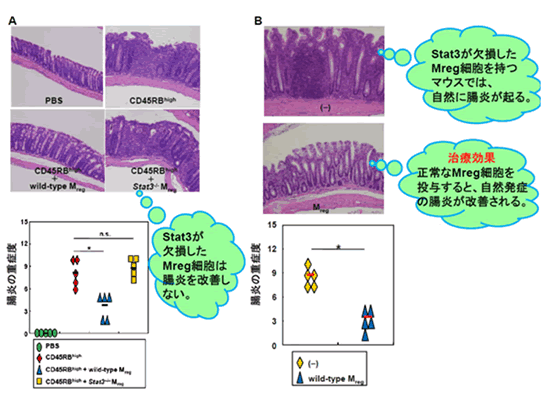
図2 Stat3欠損Mreg細胞は腸炎抑制能を持たない
- A IBDモデルとしてRag2 -/-マウスにCD4+ CD45high T細胞を3x105個移入。同時に、PBS、野生型Mreg細胞(3x105個)、Stat3-/-Mreg細胞(3x105個)を投与。
- B Stat3を欠損したMreg細胞を持つLysM-cre;Stat3 F/Fマウスは、腸炎を自然に発症する。正常なMreg細胞を投与すると腸炎の発症が抑制される。コントロール群にはPBS投与を行った。
本研究の結果より、①大腸に存在するMreg細胞は、細胞接着分子を高発現し炎症性T細胞と優位に結合する、②Mreg細胞では、IL-10やStat3が炎症性T細胞の活性化に必要なCD80やCD86の発現を抑制しているため、炎症性T細胞の増殖を誘導できない、という2段階の方法で炎症性T細胞の活性化を抑制し、腸管免疫系の恒常性維持に重要な役割を果たすことが推測されました。
本研究では、マウスの大腸に存在する自然免疫細胞のうちMreg細胞が、腸炎の発症を抑制していること、さらに、腸炎モデルマウスにおいてMreg細胞の腹腔への投与が治療効果を示すことを明らかにしました。今後、ヒトのMreg細胞の同定および作用機序の解明が進むことにより、IBDの効果的な治療法開発につながることが期待されます。
近年、腸内細菌叢注8)の変化に伴う腸管免疫寛容の破綻が、IBDだけでなく多発性硬化症などの自己免疫疾患の発症に深く関与することが報告されています。いずれも、欧米化された食生活をはじめとするライフスタイルの変化に伴い急激に日本で増加している難病です。ヒトMreg細胞が発見され細胞培養により多量のMreg細胞を作ることが可能となれば、IBDだけではなく多くの自己免疫疾患の治療法開発に結びつくことが期待されます。
<用語解説>
- 注1)免疫細胞
- 生体内で病原体やがん細胞を排除するために働く細胞。元来生体に備わっている自然免疫を担当する細胞(自然免疫細胞)と決まった抗原による刺激で誘導される獲得免疫を担当する細胞(獲得免疫細胞)に大別される。また、免疫細胞はもともと共通の造血幹細胞から分化するが、その後の分化する器官により、骨髄中で分化する細胞(ミエロイド細胞)と胸腺中で分化する細胞(リンパ球系細胞)に分類することもできる。
- 注2)自然免疫細胞
- 体内に病原体が侵入した際に、最初の生体防御反応として機能する免疫細胞。異物の構成成分を認識することで活性化し、病原体を貪食する。マクロファージや樹状細胞などが含まれる。
- 注3) 獲得免疫細胞
- 自然免疫細胞が貪食し、提示した病原体の特徴的分子に対して病原体に対抗する免疫反応を行う細胞でT細胞とB細胞が主体となる。
- 注4)免疫寛容
- 病原性のない微生物や自己の細胞に対して免疫細胞が反応を示さない状態。
- 注5)炎症性腸疾患(IBD)
- 免疫細胞により腸の粘膜が攻撃され炎症を起こす自己免疫疾患で、発症の原因は分かっていない。潰瘍性大腸炎やクローン病などがある。
- 注6) 細胞接着分子
- 細胞膜に存在して細胞同士の結合に関与するたんぱく質。
- 注7) 転写因子
- 遺伝子に結合して、遺伝子の発現を調節するたんぱく質。
- 注8) 腸内細菌叢
- 腸内には100種類以上、100兆個以上の腸内細菌が生息しており、宿主であるヒトや動物の栄養分の一部を利用しながら、他の腸内細菌とバランスを取って一種の生態系を形成しており、この生態系を腸内細菌叢(腸内フローラ)と呼ぶ。
<共同研究>
本研究は、大阪大学 大学院生命機能研究科の村上 正晃 准教授、大阪大学 免疫学フロンティア研究センターの石井 優 教授、徳島大学 疾患ゲノム研究センターの岡崎 拓 教授、順天堂大学 医学部の八木田 秀雄 専任准教授の協力を得て行われました。
<原論文情報>
“Intestinal CX3C chemokine receptor 1high (CX3CR31high) myeloid cells prevent T-cell-dependent colitis” Proc. Natl. Acad. Sci. USA. 109, 5010 (2012).
(腸管粘膜固有層に存在するCX3CR1highミエロイド細胞はT細胞依存的な腸炎を抑える)
Proc Natl Acad Sci U S A. 2012 Mar 27;109(13):5010-5. doi: 10.1073/pnas.1114931109. Epub 2012 Mar 7.
Kayama H1, Ueda Y, Sawa Y, Jeon SG, Ma JS, Okumura R, Kubo A, Ishii M, Okazaki T, Murakami M, Yamamoto M, Yagita H, Takeda K.