
3.抗原提示のしくみ
体外から侵入してきた病原体等の抗原蛋白質は抗原提示細胞である樹状細胞によって積極的に細胞内へ取り込まれます。また、B細胞では、細胞膜上に発現する抗体分子が抗原受容体(B細胞受容体:BCR)として働き、抗原蛋白質を細胞内へ取り込みます。これら細胞内に取り込まれた抗原蛋白質は抗原ペプチドに分解された後で、主要組織適合抗原(MHC)注1) クラスII分子によって細胞表面に抗原提示されます。T細胞は、これらMHCクラスII分子によって提示された抗原をT細胞受容体(TCR)によって認識し、ヘルパーT細胞へと分化します。ヘルパーT細胞のヘルプを受けて、B細胞は強い抗体産生細胞へと分化します。他方、細胞内で産生される抗原蛋白質(例えばウイルス蛋白質等)は、MHCクラスI分子によって細胞表面に抗原提示されます。キラーT細胞は、これらMHCクラスI分子によって提示された抗原を認識し、細胞を傷害します。このように獲得免疫の応答は、MHC分子による抗原ペプチドの抗原提示から始まります。一方、MHCの遺伝子多型は自己免疫疾患の感受性を最も強く左右することが知られており、どのように特定のMHCが自己免疫疾患の感受性に関与するかを明らかにすることは、自己免疫疾患の原因や治療法を開発するうえで非常に重要であります。しかし、依然として特定のMHCがどのように自己免疫疾患に関与するかは明らかになっておりませんでした。
荒瀬チームは、細胞内で産生された自己抗原蛋白質が異常に折り畳まれる(ミスフォールドされる)と、いままで自己免疫疾患に感受性と言われてきたMHCクラスII分子と会合できるようになり、それらが細胞表面に運ばれて、自己抗原として、B細胞へ抗原提示されることを明らかにしました (Jiang et al. Int. Immunol 2013)。さらに、この新たなミスフォールド蛋白の抗原提示が自己免疫疾患の発症要因である自己抗体産生に関わるという新たな仮説を提唱しています (Jin et al. Proc. Natl. Acad. Sci. USA 2014)。
【荒瀬チーム】「関節リウマチ等の自己免疫疾患の新たな発症機構を発見」PNAS 2014
―自己免疫疾患の診断薬・治療薬開発へ繋がる新たな分子機構―
<ポイント>
- MHCクラス II分子が細胞内の変性タンパク質を細胞外へ輸送することを発見した。
- MHC クラスII分子によって細胞外へ輸送された自己のミスフォールドタンパク質が自己免疫疾患の自己抗体の標的分子であることを明らかにした。
- MHCクラスII分子によるミスフォールドタンパク質の輸送能が、MHCクラスII分子の自己免疫疾患感受性と高い相関を示すことを明らかにし、変性タンパク質/MHCクラスII分子複合体が自己免疫疾患の発症に関わっていることを明らかにした。
自己免疫疾患は、自己分子に対する抗体(自己抗体)等が自己組織を誤って攻撃してしまうことで生じる疾患です。しかし、なぜ自己免疫疾患で自己抗体が産生されるかは、依然として明らかでありません。MHCは、細胞内外のタンパク質が細胞内でペプチドに分解されたものを細胞表面に輸送してT細胞に抗原として提示することで、免疫応答の中心を担っています。一方で、MHCは、自己免疫疾患の罹りやすさに最も影響を与える原因遺伝子として知られていますが、MHCがどのように自己免疫疾患を引き起こすかも明らかでありませんでした。
本研究では、通常は速やかに分解されてしまう細胞内の変性タンパク質が、MHCクラスII分子によって細胞外へ誤って輸送されてしまい、その変性タンパク質が自己抗体の標的分子であることを世界で初めて明らかにしました。つまり、MHCクラスII分子が細胞内の変性タンパク質を自己応答性のB細胞に提示することが自己免疫疾患の原因であると考えられました。実際に、関節リウマチ患者の血液を解析すると、MHCクラスII分子によって細胞外へ運ばれた変性タンパク質に対する特異的な自己抗体が認められることが判明しました。さらに、変性タンパク質と結合しやすいMHCクラスII分子を持っているヒトは持っていないヒトに比べて10倍以上も関節リウマチになりやすいことを発見しました。これらの結果から、MHCクラスII分子によって細胞外へ輸送されてしまった細胞内の変性タンパク質が、自己免疫疾患の発症に関与していることが判明しました。
本研究によって、今まで考えられてきた自己免疫疾患の発症機序(図1)とは全く異なる新たな発症機序が明らかになり、自己免疫疾患でなぜ自己抗体が産生されるのか、なぜ主要組織抗原が自己免疫疾患に関わっているかという長年の自己免疫疾患の謎を解明する上で、非常に重要な発見です。関節リウマチに限らず、その他多くの自己免疫疾患も同様な発症メカニズムが考えられるため、本研究成果は、今後、多くの自己免疫疾患の治療薬や診断薬の開発に貢献することが期待されます。
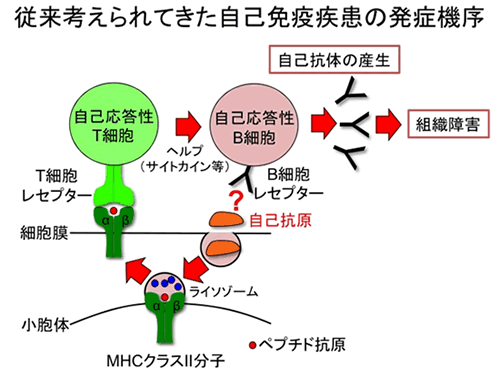
図1 従来考えられてきた自己免疫疾患の発症機序
自己免疫疾患は、自己に対する抗体等が自己組織を誤って攻撃してしまうことで生じる疾患です。主要組織適合抗原(MHC)は、非常に多様性に富む分子で、それぞれの個人で異なる組み合わせを持っており、どのMHCを持っているかで、自己免疫疾患の感受性が決定される最も重要な分子です。MHCはペプチド抗原をT細胞に提示することから、自己免疫疾患の原因はT細胞の異常だと長年考えられてきましたが、依然として、自己免疫疾患の原因は明らかになっていません(図1)。
関節リウマチは、免疫機構が関節を破壊してしまう自己免疫疾患で、人口の約1%が罹患すると言われている最も頻度の高い代表的な自己免疫疾患です。関節リウマチ患者の血液には、様々な自己抗体が認められます。自己抗体は関節リウマチの発症に直接関与している一方、関節リウマチの診断にも使われています。リウマトイド因子注2)は、変性した抗体に対する自己抗体であり、関節リウマチ患者の約8割が陽性であることから、50年以上前から関節リウマチの診断に使われています。しかし、関節症状のない他の自己免疫疾患および正常人でも陽性になることがあります。また、変性した抗体は生体内に存在しないため、リウマトイド因子が本来何を認識する自己抗体なのか、なぜ関節リウマチで陽性になるかは不明でした。
一方、細胞内では正常蛋白質ばかりでなく、うまく折りたためられなかった変性タンパク質も常に作られています。しかし、そのような変性タンパク質は細胞内で速やかに分解されてしまい細胞外に運ばれることはありません。ところが、本研究によって細胞内の変性蛋白質が自己免疫疾患に感受性のMHCクラスII分子と結合すると、変性タンパク質がMHCクラスII分子によって細胞外に輸送され、それが異物として自己抗体の標的になることが判明しました(図2)。
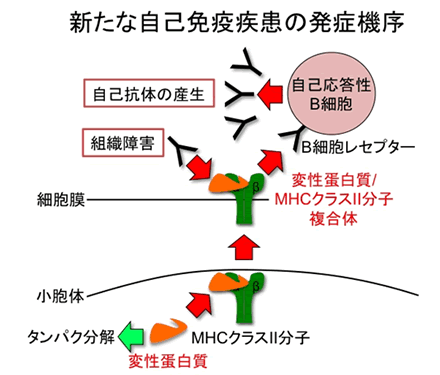
図2 今回明らかになった新たな自己免疫疾患の発症機序
MHCクラスII分子がリウマトイド因子の変性抗体の認識に関わっているかを調べるために、ヒト抗体重鎖遺伝子と共にヒトMHCクラスII遺伝子をヒト細胞に遺伝子導入しました。抗体は軽鎖と重鎖から成るため、重鎖のみでは変性して細胞外に輸送されることはありません。ところが、MHCが存在すると、抗体重鎖がMHCクラスII分子と結合して細胞表面に出現することが判明しました。さらに、この変性抗体重鎖とMHCクラスII分子複合体は関節リウマチ患者血液中の自己抗体に認識されることが判明しました(図3)。さらに多くの関節リウマチ患者の血液を調べてみると、今まで診断に使われてきたリウマトイド因子の値と変性抗体/MHCクラスII分子複合体に対する抗体量は強く相関しました(図4)。ところが、関節症状のない他の自己免疫疾患および正常人血清を解析してみると、リウマトイド因子陽性の血液でも変性抗体/MHCクラスII分子複合体に対する抗体は認められませんでした。以上より、今まで診断に使われてきたリウマトイド因子と比べて、変性抗体/MHCクラスII分子複合体は、関節リウマチ患者に特異的な自己抗体の標的であることが判明しました。
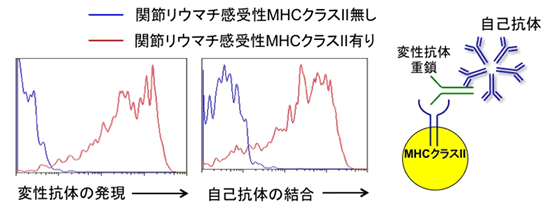
図3 関節リウマチ患者の自己抗体は、MHCクラスII分子によって細胞外へ輸送された変性抗体重鎖を認識する。
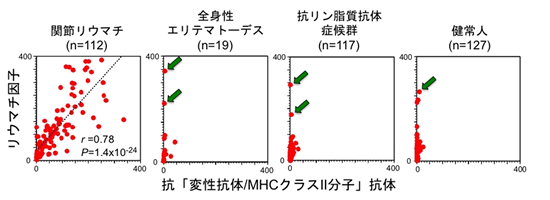
図4 変性抗体/MHCクラスII分子複合体は、関節リウマチで産生される自己抗体の特異的な標的分子である。
次に変性抗体/MHCクラスII分子複合体が、実際に関節リウマチ患者の関節滑膜に存在するかどうかを関節リウマチ患者の滑膜組織を用いてPLA法注3)で解析しました。その結果、変性抗体/MHCクラスII分子複合体が関節リウマチ患者の関節滑膜に存在するが、自己免疫疾患ではない変形性関節症の患者の関節滑膜には存在しないことが判明しました(図5)。従って、変性抗体/MHCクラスII分子複合体が関節リウマチ患者の関節滑膜で産生され、それが自己抗体の標的になっていると考えられました。
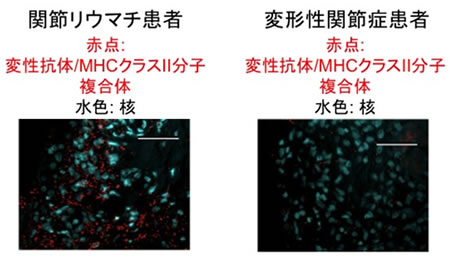
図5 変性抗体/MHCクラスII分子複合体が、関節リウマチ患者の関節滑膜に認められる。
最後に、変性抗体/MHCクラスII分子複合体が関節リウマチの発症に関わっているかを調べました。関節リウマチの罹りやすさはMHCクラスIIの型(アリル)によって決定されることが知られています。例えばヒトMHCクラスIIの一つであるHLA-DR4を持っているヒトは、HLA-DR3を持っているヒトより約10倍以上も関節リウマチに罹りやすくなります。そこで、抗体重鎖と種々のHLA-DRとの複合体に対する自己抗体の結合性を解析しました。その結果、それぞれのHLA-DRを持っているヒトの関節リウマチの罹りやすさ(オッズ比)と変性抗体/HLA-DR複合体に対する自己抗体の結合性は、非常に高い相関を示すことが判明しました(相関係数0.81、危険率0.000046)(図6)。つまり、関節リウマチに罹りやすいMHCを持っているヒトは、自己抗体の標的抗原が産生されやすいことになります。以上の結果より、変性抗体/MHCクラスII分子複合体が自己抗体の標的として関節リウマチの発症に関わっていると考えられました。
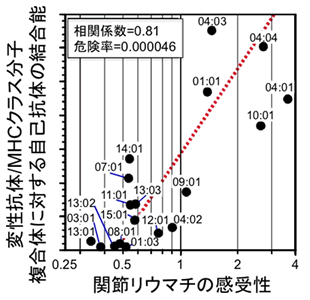
図6 変性抗体/MHCクラスII分子複合体に対する自己抗体の結合は関節リウマチの感受性(罹りやすさ)と強い相関を示す。
本研究により、変性蛋白質とMHCとの分子複合体が自己抗体の標的として、関節リウマチの発症に関わっていることが明らかになりました。他の自己免疫疾患においても同様に変性蛋白質とMHCとの複合体が自己抗体の標的になっていると思われます(論文投稿中)。従って、変性蛋白質/MHCクラスII分子複合体は様々な自己免疫疾患の治療薬開発のための標的分子だと思われます。また、変性蛋白質/MHCクラスII分子複合体に特異的な自己抗体が産生されることから、MHCと変性蛋白質との複合体は自己抗体の検出にも有用であり、自己免疫疾患の診断にも役立ちます。今後、様々な自己免疫疾患での変性蛋白質/MHCクラスII分子複合体の研究を進めることによって、自己免疫疾患の病因解明が期待されます。
<用語解説>
- 注1)主要組織適合抗原(Major Histocompatibility Complex, MHC; Human Leukocyte Antigen, HLA)
- MHCは非常に多様性に富む分子であり、基本的に全てのヒトが異なるMHCを持っている。T細胞にペプチド抗原を提示する(図2)ことで、免疫応答の中心を担っている分子である。クラスIとクラスIIがあり、クラスIIはヘルパーT細胞に抗原を提示することで、B細胞の抗体産生に関与していると考えられている。また、ヒトのクラスIIはHLA-DRとも呼ばれている。一方、MHCは、以前より自己免疫疾患の発症に最も関与した分子であることが知られており、最近の全ゲノム解析によっても、主要組織抗原が最も強く自己免疫疾患の感受性に関与した遺伝子であることが確認された。しかし、なぜ特定のMHCを持っていると特定の自己免疫疾患になりやすいかは、依然として明らかになっていなかった。
MHC分子の今まで考えられていた特徴
MHCクラスI分子 MHCクラスII分子 抗原ペプチドの由来 細胞内由来タンパク質(ウイルスなど) 細胞外由来タンパク質(細菌など) ペプチド生成機構 プロテアソーム エンドソーム 発現細胞 全ての細胞 免疫系細胞 インバリアント鎖の有無 なし あり T細胞への結合 キラーT細胞の持つCD8と結合 ヘルパーT細胞の持つCD4と結合 - 注2)リウマトイド因子
- 最も昔から知られている自己抗体の一つであり、変性した抗体に対する自己抗体である。約8割の関節リウマチの患者で陽性になり、現在でも関節リウマチの検査に使われている。しかし、関節症状のない他の疾患や健常人でも陽性になることがある。しかし、変性した抗体は通常生体内に存在しないため、どのような抗原がリウマトイド因子を誘導するのか、なぜ、関節リウマチの陽性率が高くなるのかが明らかになっていない。
- 注3)PLA法 (Proximity Ligation Assay)
- 組織や細胞内での分子間相互作用を検出する方法。40nm以下の分子間の近接を検出することができる。
<原論文情報>
- Yan Jiang, Noriko Arase, Masako Kohyama, Kouyuki Hirayasu, Tadahiro Suenaga, Hui Jin, Maki Matsumoto, Kyoko Shida, Lewis L. Lanier, Takashi Saito and Hisashi Arase
Transport of misfolded endoplasmic reticulum proteins to the cell surface by MHC class II molecules.
Int. Immunol. 25:235-246 2013. - Hui Jin, Noriko Arase, Kouyuki Hirayasu, Masako Kohyama, Tadahiro Suenaga, Fumiji Saito, Kenji Tanimura, Sumiko Matsuoka, Kosuke Ebina, Kenrin Shi, Noriko Toyama-Sorimachi, Shinsuke Yasuda, Tetsuya Horita, Ryosuke Hiwa, Kiyoshi Takasugi, Koichiro Ohmura, Hideki Yoshikawa, Takashi Saito, Tatsuya Atsumi, Takehiko Sasazuki, Ichiro Katayama, Lewis L. Lanier, and Hisashi Arase
Autoantibodies to IgG/HLA class II complexes are associated with rheumatoid arthritis susceptibility.
Proc. Natl. Acad. Sci. USA. 111:3787-3792 2014.