柳澤 修一(東京大学 大学院農学生命科学研究科 助教授)
栄養シグナルによる植物代謝制御の分子基盤
平成17年度 平成18年度 平成19年度
§1.研究実施の概要
炭酸ガスや硝酸イオンといった無機物は植物栄養として生体物質の合成に使われるだけでなく、遺伝子発現の制御や代謝の調節に関わるシグナルとしてはたらき、植物栄養は植物の生長や物質の生産量に深く関与している。また、無機物から生合成される初期産物である糖やアミノ酸なども、エネルギー源あるいは他の有機物の生合成の原料となると同時に栄養シグナル伝達物質として働いている。植物は、個々の栄養シグナルによる制御のみならず、様々な栄養シグナルの制御を総合して植物栄養を効率よく利用し、生長と物質生産を行っていると考えられる。植物栄養に応答した遺伝子発現の制御と代謝調節の仕組みを明らかにすることは植物の物質生産能力を活用していく上で非常に重要であるにもかかわらず、現在のところ、そのごく一端がわかっているのみである。そこで、植物栄養による生長・物質生産の制御の包括的理解を目指して、栄養シグナルの伝達の仕組みと複数の栄養シグナルに基づく遺伝子発現と代謝の制御の仕組みの解明を目指す。特に、植物における物質生産の中心である炭素と窒素の同化と同化のためのエネルギー供給システムに着目し、これらの代謝経路が相互に影響を及ぼしながら植物栄養を効率よく同化して物質生産を行っている仕組みの解明に力点を置く。我々は、遺伝子導入による代謝改変に成功しており、ラン藻の酵素の遺伝子の導入による炭素代謝能改変植物体、転写因子の遺伝子の導入による窒素同化能力強化植物、NADキナーゼ過剰発現によるエネルギーバランス改変植物の作出を行っている。今年度までに、これら形質転換植物を用いたメタボローム解析や包括的遺伝子発現解析などを行い、個々の代謝改変の分子的基盤を明らかにしつつある。次年度以降は、代謝改変を引き起こす遺伝子の多重導入によって今までに得られた知見を更に評価する予定である。栄養シグナルの伝達の仕組みに関しては、今年度までに植物栄養シグナルの伝達にはタンパク質のリン酸化や分解の制御が重要であることを示すデータを取得し、それに基づき栄養応答に関わるタンパク質の変化を包括的に解明するための植物タンパク質のプロテオーム解析方法を確立した。次年度以降は、これまでに単離している植物栄養シグナル応答変異株などを用いたプロテオーム解析により栄養シグナルの伝達と応答を総合的に解析する予定である。
§2.研究実施内容
植物栄養による生長と物質生産量を制御する仕組みの解明を目的として、炭素代謝能改変植物、窒素同化能力強化植物、エネルギーバランス改変植物を用いたメタボローム解析や包括的遺伝子発現解析などを実施した。 窒素同化に必要な炭素骨格の生合成経路の酵素の発現を促進するトウモロコシ転写因子Dof1を発現しているシロイヌナズナ形質転換体や形質転換ジャガイモでは、遊離アミノ酸の総量の増加が見られることを昨年度までに示している。今年度は、シロイヌナズナ形質転換体の詳細なメタボローム解析を行い、アンモニウムイオン非存在下で生育させた場合には地上部と根のいずれにおいてもDof1形質転換体とコントロール植物体の遊離アミノ酸含量の間に有意な相違は確認されないが、アンモニウムイオン存在下で生育させた場合には地上部でのみコントロール植物体に比べてDof1形質転換体のほうが高い遊離アミノ酸含量を持つこと、また、アンモニウムイオン存在下で生育させた形質転換体の地上部でのみ遊離アミノ酸の含量の増加ともに窒素同化に必要な炭素骨格の供給に関わるクエン酸回路の代謝中間体であるリンゴ酸およびフマル酸の減少も観察されることを見いだした。この現象を利用して、培地中のアンモニア濃度を変化させることによりDof1によって直接的に改変された代謝経路とその代謝改変に基づく付随的な代謝変化を調べ、異なる同化経路間の相互作用を評価した。また、同時にDNAマイクロアレイ解析を行うことにより、遺伝子の発現レベルでも同様の評価を行った。その結果、図1に示すように、Dof1単独では硝酸からアンモニアへの還元段階には影響を及ぼさず窒素同化のための炭素骨格供給経路のみを弱く活性化しているだけでありが、この状態にアンモニアが供給されると窒素の同化が強力に進み、これにより炭素ガスの同化が促進されさらに炭素骨格の供給が増し、窒素同化がさらに促進されるという正の制御のループが形成されていると見られた。さらに、増強された窒素の同化はアミノ酸プールを大きくすると同時に硫黄の同化に必要な多数の遺伝子の発現を促進することを見いだした。硫黄の同化はセリンとアセチル-CoAからO-アセチルセリンが合成されることが必須であるが、実際、Dof1形質転換体ではアンモニウムイオン依存的にO-アセチルセリンと代表的な硫黄同化物であるグルタチオンのレベルの上昇が確認された。したがって、窒素同化の強化は副次的に炭素同化と硫黄同化をも増強し(図1参照)、炭酸固定経路と窒素同化経路
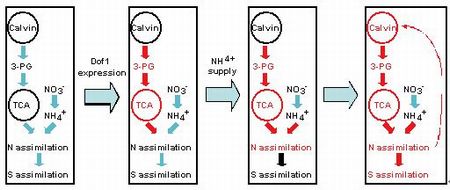
図1.Dof 1形質転換体における代謝改変の概念図
の相互依存的活性化とこの二つの同化経路の活性化による硫黄同化に関わる遺伝子群の発現の促進および硫黄同化の促進が判明した。このことから、今後、Dof1形質転換体の解析から硫黄同化の鍵を握る転写因子の同定も可能であることが示唆された。このようなDof1の効果は光合成を強化した場合により大きくなると期待される。そこで、現在 、Dof1と同時に、葉緑体で発現させた場合に光合成同化能力の強化を導くラン藻の酵素、フルクトース-1,6-/セドヘプツロース-1,7-ビスホスファターゼ (FBP/SBPase)を発現している形質転換体の作出を試みている。
Dof1形質転換体の解析に加え、今年度は、FBP/SBPaseを細胞質で発現しているタバコ(TcFS)の代謝変化とエネルギー代謝改変植物における炭素と窒素代謝の変動の評価も行った。通常CO2環境下(360 ppm)で栽培したTcFSの光合成能および生育は野生株のものと有意な相違は認められなかったが、光合成機能向上が見込まれる高CO2環境下(1200 ppm)での両株の生育を比較した場合、TcFSでは側枝、葉数、茎径、乾燥重量のいずれも野生株と比較してさらに増大しており、また、野生株ではソース葉にヘキソース類の蓄積していたのに対し、TcFSのソース葉ではヘキソース類の蓄積が認められずシンク葉でのショ糖およびデンプンの蓄積が増大していた。このことから、細胞質FBPaseは光合成活性が上昇する条件下ではショ糖合成系の律速となり、また、ヘキソースなどの代謝中間体が形態形成のシグナルとなる可能性が示唆された。一方、植物に特異的なプラスチド局在型NADリン酸化酵素(NADK2)の過剰発現体、および欠損植物体における光合成能力、アミノ酸合成能力、およびその代謝産物の詳細な定量実験により、エネルギー代謝改変植物おける炭素と窒素代謝の変動を評価した。NADK2の過剰発現体では葉の緑色が濃く生育が若干早くなる一方、欠損植物体では葉の色は薄黄緑となり生育も遅延した。パルス変調を用いた蛍光測定による光化学系の活性の比較から生育増進の原因が光合成の光化学系の変化によるものでは無いことが示されたので、カルビン回路とその周辺の代謝物の分析を行った。過剰発現体では炭酸固定の対象であるリブロース1,5-2リン酸やアミノ酸の炭素骨格となる2-オキソグルタル酸の増加が検出され、また、調べたアミノ酸の多くが過剰発現体で増加していた。さらに、カルビン回路の中でも特に重要と考えられる酵素の活性を比較した結果、CO2を固定する反応を触媒するRuBisCO、及びカルビン回路内で唯一NADPH依存性の酵素であるGAPDH、また、還元型チオレドキシンにより活性化することが知られているFBPaseのいずれにおいても過剰発現体での酵素活性が上昇していた。これらの結果から、エネルギー代謝の改変により、炭素と窒素の双方の代謝増進が起きたことが示唆された。エネルギー代謝の改変と炭素と窒素の同化能力の関係を精査するために、現在、NAD合成に関与する酵素の過剰発現シロイヌナズナ及びNAD合成に関与する酵素とNADK2の両方を過剰発現している2重形質転換体の作出を行っている。
栄養シグナル伝達の仕組みを明らかにするためには、構成因子を同定することが極めて重要である。CO2濃度変化に伴う気孔開度の変化とそれに基づく葉表面温度の変化を指標にCO2応答性突然変異体を探索し、低CO2条件下でも高温を示す変異体としてht1(high leaf temperature 1)変異体を同定して、炭酸ガスが重要な植物栄養シグナル(炭素シグナル)として機能していることを昨年度までに示している。今年度は、ht1変異体のスクリーニングに用いた類似の方法により、CO2非感受性シロイヌナズナ変異株cdi (cdi1-cdi11)を単離し、現在、原因遺伝子の同定を行っている。また、炭素シグナルと窒素シグナルの伝達あるはそれらの応答に関わる因子を探索するために、新たなスクリーニング方法を確立して変異株を単離した。すなわち、アクティベーションタグラインを用いてグルタミン合成酵素阻害剤であるメチオニンスルホキシミン(MSX)存在下で異なる表現型を示す変異体のスクリーニングを行い、有意にMSX耐性を示す株を3株、野生株よりもMSXに対して高い感受性を示す株を5株単離した。これらの変異株についても、現在、原因遺伝子の同定を行っている。一方で、HT1はタンパク質リン酸化酵素であったことや、炭酸ガスから生合成される糖はある種の転写因子の分解を促進することを明らかにしていることから、植物栄養シグナル伝達および応答の仕組みを解明するうえでタンパク質の包括的解析が重要であると判断した。そこで、今年度は、植物タンパク質のプロテオーム解析の系の確立を行った。ゲル電気泳動、イオン交換クロマトグラフィー、限外濾過の3種類の分画方法を用いてタンパク質とペプチドを分画し、ナノフロー液体クロマトグラフィーとオンラインで接続されたイオントラップ型質量分析計を用いてタンパク質を同定するシステムを確立した。まず、最初に、植物栄養応答の観点から重要であり、かつ、存在するタンパク質の種類が比較的少ないことが予測された高等植物における物質運搬システムである導管と篩管に存在するタンパク質の同定を行った。この結果、導管液と篩管液には100種類強のタンパク質と数種類のペプチドが存在することを明らかにした。従来から篩管液中に存在が知られていた酸化還元や生体防御反応等に関与するタンパク質に加え、ホメオドメイン型やジンクフィンガー型の転写因子と推定されるタンパク質も同定できたことから、篩管は単なる物質輸送器官ではなくシグナルの伝達においても重要な役割を果たしていることを示唆した。さらに、今年度は、核タンパク質の同定も開始し、現在までに、400種程度のタンパク質を同定している。半定量的に同定する方法も確立したので、次年度は植物栄養に応答したタンパク質の変動を定量的かつ包括的に解析し、栄養シグナル伝達あるいは応答に関わる因子の同定を行う予定である。
§3.成果発表等
論文(原著論文)発表
(1)発表総数(国内 0件、国際 11件)
(2)論文詳細情報
- Yoshinaga, K., Fujimoto, M., Arimura, S.I., Tsutsumi, N., Uchimiya, H., Kawai-Yamada, M. (2006) Mitochondrial fission regulator, DRP3B does not regulate cell death in plant. Ann. Bot. 97, 1145-1149.
- Takahashi, H., Hayashi, M., Goto, F., Sato, S., Soga, T., Nishioka, T., Tomita, M., Kawai-Yamada, M., Uchimiya, H. (2006) Evaluation of metabolic alteration in transgenic rice overexpressing dihydroflavonol-4-reductase. Ann. Bot., 98, 819-825
- Ogawa, T., Uchimiya, H., Kawai-Yamada, M. (2006) APETALA2, a plant floral homeotic gene, is regulated by Arabidopsis thaliana ethylene-responsive element binding protein in Arabidopsis plant. Ann Bot., 99, 239-244.
- Takahashi, H., Watanabe, A., Tanaka, A., Hashida, S., Kawai-Yamada, M., Sonoike, K., Uchimiya, H. (2006) Chloroplast NAD kinase is essential for energy transduction through xanthophyll cycle in photosynthesis. Plant Cell Phys., 47, 1678-1682.
- Hashida, S., Takahashi, H., Kawai-Yamada, M., Uchimiya, H. (2007) Arabidopsis thaliana nicotinate/nicotinamide mononucleotide adenyltransferase (AtNMNAT) is required for the growth of pollen tube. Plant J. 49, 694-703
- Ihara-Ohori, Y., Nagano, M., Muto, S., Uchimiya, H., Kawai-Yamada, M. (2007) Cell death suppressor, Arabidopsis BI-1, is associated with calmodulin-binding and ion homeostasis. Plant Phys., 143, 650-660.
- Kusumi, K., Yaeno, T., Kojo, K., Hirayama, M., Hirokawa, D., Yara, A. and Iba, K. (2006) The role of salicylic acid in the glutathione-mediated protection against photo-oxidative stress in rice. Physiol. Plant 128: 651-661.
- Kojo, K., Yaeno, T., Kusumi, K., Matsumura, H., Fujisawa, S., Terauchi, R. and Iba, K. (2006) Regulatory mechanisms of ROI generation are affected by rice spl mutations. Plant Cell Physiol. 47: 1035-1044.
- Hamada, T., Iba, K. and Shimada, T. (2006) Reduction of trienoic fatty acid content by expression of a double-stranded RNA of a plastid ω-3 fatty acid desaturase gene in transgenic tobacco. Biotechnol. Lett. 28: 779-785.
- Hashimoto, M., Negi, J., Young, J., Israelsson, M., Schroeder, J.I. and Iba, K. (2006) Arabidopsis HT1 kinase controls stomatal movements in response to CO2. Nature Cell Biol. 8: 391-397.
- Yaeno, T., Saito, B., Katsuki, T. and Iba, K. (2006) Ozone-Induced expression of Arabidopsis FAD7 gene requires salicylic acid, but not NPR1 and SID2. Plant Cell Physiol. 47: 355-362.
![]()