JST 課題解決型基礎研究の一環として、曽我部 正博 名古屋大学 大学院医学系研究科 教授らの研究グループは、運動する鞭毛・繊毛注1)が力を感じる能力(機械受容能)も持ち合わせていることを初めて明らかにしました。
鞭毛・繊毛は、精子や気管上皮細胞などでは運動器官としてのみ働き、一方、腎臓などにある一次繊毛注2)は運動機能を持たず、機械刺激注3)を感知するといった感覚器官としての機能のみを持つことが知られています。このように、従来、鞭毛・繊毛は運動器官として、または感覚器官として機能するものが別々に2種類あると考えられてきましたが、本研究で初めて、運動機能と感覚機能を併せ持つ鞭毛・繊毛があることが明らかになりました。本研究で用いた単細胞真核生物のクラミドモナスは、細胞の前端にある鞭毛を用いて遊泳運動をしていますが、障害物に衝突した時は一時的に後退遊泳をします。クラミドモナスは、衝突を鞭毛の変形として鞭毛の根元で感じ、細胞の電気的活動を引き起こすことを明らかにしました。すなわち、クラミドモナスの鞭毛は運動器官として働くだけではなく、力による変形を感じる感覚器官としての機能を兼ね備えているということが分かりました。さらに、力による変形を実際に受け取る分子実体は、多種の刺激を受け取るたんぱく質として知られているTRPチャネル注4)の一種であることを突き止めました。
鞭毛・繊毛の運動機能の異常が内臓逆位や不妊の遺伝病の原因となり、感覚機能の異常が多発性嚢胞腎注5)の原因となるなど、鞭毛・繊毛の機能はヒトの疾病と密接な関連があることが分かっています。
本研究により、鞭毛・繊毛にかかる負荷を鞭毛・繊毛自身が検出し、運動を制御しているというフィードバックシステム注6)があり、運動機能と感覚機能が連携していることが分かりました。この成果は、運動する鞭毛・繊毛による機械受容能と運動制御の研究の突破口となり、鞭毛・繊毛に関係する疾病の解明につながると期待されます。
本研究は、筑波大学、東京学芸大学と共同で行ったもので、本研究成果は、2011年4月10日(英国時間)に英国科学雑誌「Nature Cell Biology」のオンライン速報版で公開されます。
本成果は、以下の事業・研究プロジェクトによって得られました。
戦略的創造研究推進事業 発展研究(ICORP-SORST)
事業・研究プロジェクト概要
| 研究プロジェクト |
「ナノ・マイクロ超分子複合体によるメカノトランスダクション機構の解明」 |
| 研究総括 |
曽我部 正博(名古屋大学 大学院医学系研究科 教授) |
| 研究期間 |
平成17年1月~平成22年3月 |
JSTはこのプロジェクトで、細胞が環境・細胞の成長・分裂・形態変化・運動に由来する機械刺激をどのように受容するかを解明し、基礎生物学から臨床医学に貢献する研究を行っています。
<研究の背景と経緯>
真核生物の鞭毛・繊毛は、運動器官としての機能は以前から知られていましたが、最近、機械刺激を感知するなど感覚機能を担う器官として注目が集まっています。例えば、腎臓の尿細管の一次繊毛は、尿細管の流速を測定する感覚器であり、この感覚の異常は多発性嚢胞腎という重篤な遺伝病の原因となっていることが明らかになってきました。一次繊毛は、ヒトの多くの器官にある運動機能を持たない繊毛で、さまざまな感覚機能を担っていると推定されています。鞭毛・繊毛がどこでどのような感覚を担っているかは、ヒトの器官の働きと疾病を解明する上で、重要な研究テーマです。
従来、鞭毛・繊毛は運動機能のみを有し感覚機能を持たないタイプと、これとは逆に感覚機能のみを有し運動機能を持たないタイプの2種類があると考えられてきました。一方、表面に繊毛が密に生えた気管繊毛上皮注7)の繊毛の運動が、粘膜の負荷が上昇すると活発になったり、ゾウリムシのような繊毛虫が障害物に衝突すると繊毛の打ち方が逆転し後退遊泳するなどという知見から考えると、鞭毛・繊毛の運動は機械刺激により制御されていると思われます。しかし、運動機能のみを持つと考えられていた鞭毛・繊毛が機械刺激を感知する感覚機能も本当に持つのか、そして、機械刺激の受容体がどこにあるのか、さらに、機械刺激受容体の分子が何であるのかは分かっていませんでした。もし、運動機能を示す鞭毛・繊毛にも感覚機能があるとしたら、鞭毛・繊毛は今まで考えられていたよりもはるかに多機能であり、生体にとって重要な役割を果たしていることになります。
そこで本研究グループでは、鞭毛の運動の実験材料として優れている単細胞生物のクラミドモナス(図1)を用いて、鞭毛に機械受容の機能があるかどうかを明らかにし、さらに、その感覚機能の分子を同定する研究に挑みました。
<研究の内容>
単細胞真核生物の一種であるクラミドモナスの鞭毛は、構造や構成たんぱく質がヒトの鞭毛・繊毛と共通性が高く、今までの研究では数多くのヒト遺伝病の原因を明らかにしてきました(図1)。また、ゲノム解析が終了し遺伝子情報が豊富に得られるなど、実験材料としての研究が進んでいます。
クラミドモナスは、前端にある2本の鞭毛を打って前進遊泳していますが、ガラス表面などの障害物に衝突すると後退遊泳をする「回避反応」を示すことが知られています(図2)。私たちは、まず、この「回避反応」がどのように生じるかということを検討するために、機械受容、温度受容、化学受容などさまざまな感覚を担うことが知られているTRPチャネルの一種が衝突を検出する機械受容の受容体ではないかという仮説を立てました。そして、TRPチャネルの阻害剤をクラミドモナスに与えたところ、回避反応が阻害されることを発見しました。さらに、ゲノム情報をもとにクラミドモナスのゲノムにあるTRPチャネルを推定したところ、7個のTRPチャネル遺伝子があることが分かりました。
クラミドモナスは鞭毛を前にして遊泳しているので、衝突すると鞭毛が変形し、それを鞭毛自身が機械刺激として受け取ると推測されます。そこで、鞭毛を脱落させた時におけるTRPチャネル発現量の変化や、抗体による局在解析の結果、少なくとも4種類のTRPチャネルが鞭毛に発現していることを明らかにしました。その中でも、TRPチャネルの一種であるTRP11は、鞭毛の根元に特に多く存在するという特徴的な分布を示していました(図3)。鞭毛は、屈曲運動をして遊泳の推進力を発生しているので、屈曲が起こるところで機械受容を行っているとは考えられません。鞭毛の根元は屈曲運動が最も小さく、しかも、衝突の時には変形しやすい位置なので、衝突を検出する機械受容の場所としては最適と考えられます。この理由から、TRP11が鞭毛で機械受容を行う受容体ではないかと推定し、RNA干渉注8)によってTRP11の発現を抑制する実験を行いました。その結果、TRP11の発現が低下した細胞では、衝突時の回避反応が見られなくなることが判明し、TRP11が鞭毛で機械受容を行っている分子であることが明らかになりました。
機械受容を担うTRP11は、機械刺激を受けると受容器電位注9)が生じます。その結果、鞭毛にある電位依存性カルシウムチャネル注10)が活性化し、細胞外から鞭毛の中にカルシウムイオンが流入すると考えられます。これが合図となってクラミドモナスの泳動方向に変化が生じる、つまり回避反応が見られるのです。
また、鞭毛に発現している電位依存性カルシウムチャネルは鞭毛の根元を避けるように鞭毛で分布していることが明らかになっています(図3)。実は、クラミドモナスの鞭毛の根元でカルシウムイオンの濃度が上昇すると、鞭毛が抜け落ちてしまうことが知られています。電位依存性カルシウムチャネルが根元に分布していないのは、回避反応を示す時に鞭毛が抜け落ちることを防ぐためだと考えられます。すなわち、長さ約15マイクロメートル(1マイクロメートル:100万分の1メートル)という微細な構造でありながら、機械受容体(TRP11)の鞭毛の根元への局在と、電位依存性カルシウムチャネル(CAV2)の鞭毛の根元以外への局在という巧妙な分布パターンによって、機械受容と鞭毛波形変換という2つの現象を両立させているといえます。
気管繊毛上皮やゾウリムシなどさまざまな鞭毛・繊毛は、負荷の変動により運動が変化します。本研究により、負荷の変化は鞭毛・繊毛自体の機械刺激受容体により受け取られ、その情報をもとに運動が制御されていることが明らかになりました(図4)。従って、鞭毛・繊毛はそれ自身に備えられたフィードバックシステムにより、負荷の変化に対応することができる運動装置であるといえます。
本研究で得られた主な成果は、以下の通りです。
- (1) 単細胞真核生物のクラミドモナスの鞭毛が、機械受容能を持つことを発見しました。
- (2) 鞭毛にある機械受容体の分子の同定に成功し、TRPチャネルの一種であるTRP11であることを示しました。
- (3) 機械受容体(TRP11)の分布は鞭毛の根元に偏っており、この分布は鞭毛自体の屈曲運動の影響を避けるのに有効であると考えられます。
- (4) これらの結果により、運動機能を持つ鞭毛・繊毛が感覚機能も併せ持つことを初めて示しました。
<今後の展開>
運動機能を持つ鞭毛・繊毛が機械受容能も併せ持つという知見は、本研究で初めて証拠が得られた考えです。しかし、気管上皮細胞の繊毛は、粘膜の負荷が大きくなると運動が活発になることが知られており、このような目で見ると、ヒトのさまざまな器官で機械受容能を持つ鞭毛・繊毛が感覚機能を担っていると想像されます。さらに、クラミドモナスの鞭毛に未解析のTRPチャネルが存在することを考えると、機械受容以外の感覚も行っていると推定されます。本研究を突破口として、運動機能を持つ鞭毛・繊毛がヒトの生体内で果たしている感覚器官としての役割の解明が進むと期待されます。
<参考図>
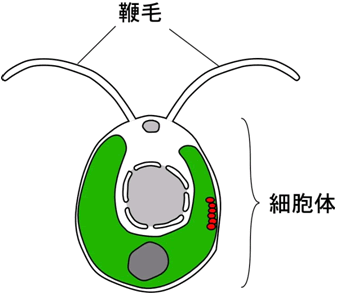
図1 クラミドモナスの細胞の模式図
細胞の前端にある2本の鞭毛を平泳ぎのように打ちながら前進遊泳する。クラミドモナスの鞭毛の構造と構成たんぱく質はほ乳類の鞭毛・繊毛と共通性が高く、鞭毛・繊毛由来の疾病の原因を数多く明らかにしてきた。鞭毛の長さは約15マイクロメートル。
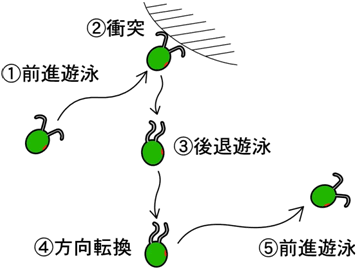
図2 クラミドモナスの回避反応
前進遊泳していた細胞が硬い障害物に衝突すると、約0.5秒間後退遊泳し、方向転換した後、新たな方向に泳ぎ去ります。衝突を感じるという機械受容により、細胞の膜電位が変化し、後退遊泳が引き起こされます。
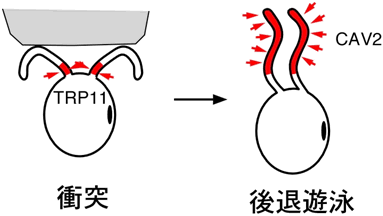
図3 クラミドモナスの鞭毛における機械刺激受容体(TRP11)と電位依存性カルシウムチャネル(CAV2)の局在
細胞が障害物に衝突すると鞭毛が折れ曲がり、根元にある機械刺激受容体(TRP11)が活性化される。その結果、細胞膜の電位が脱分極し、根元を避けるように分布する電位依存性カルシウムチャネル(CAV2)が活性化し、細胞外から鞭毛の中にカルシウムイオンが流入する。鞭毛内のカルシウムイオン濃度が上昇すると、鞭毛の打ち方が変わり、後退遊泳をする。
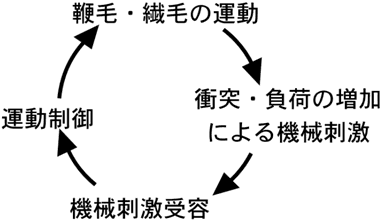
図4 鞭毛・繊毛におけるフィードバックシステムによる運動制御機構
鞭毛・繊毛運動にかかる負荷が変化すると、それが機械刺激として受容される。その結果、膜電位と細胞内カルシウムイオンの濃度が変化し、負荷の変化に対応できるように鞭毛・繊毛運動が制御される。
<用語解説>
- 注1) 鞭毛・繊毛
- 直径0.2マイクロメートル、長さ10~40マイクロメートル程度の細長い鞭状の真核生物の運動器官。精子やクラミドモナスなどのように、1、2本しかない場合は鞭毛と呼び、気管上皮細胞やゾウリムシのように多数が密生している場合は繊毛と呼ぶが、内部構造や運動機構などは同じ。原核生物の鞭毛とは異なる。
- 注2) 一次繊毛
- ほ乳類などの多細胞生物にある、運動能のない繊毛。痕跡器官と思われていたが、感覚器として重要であることが最近分かってきている。また、多くの細胞が発生の途中で一次繊毛を持つ時期があることも分かってきつつあり、注目されている。
- 注3) 機械刺激
- 変形や力といった力学的な刺激のこと。機械刺激を受容する感覚を機械受容といい、聴覚、触覚、重力感覚などが機械受容による感覚。本プロジェクトではこの細胞の感覚を、「細胞力覚」と命名している。
- 注4) TRPチャネル(Transient receptor potential cannel)
- 真核細胞にある、温度、化学刺激、機械刺激などの刺激を受容し、イオンが透過する穴を開閉するイオンチャネル。種類により、受容する刺激の種類、活性化される範囲が異なる。
- 注5) 多発性嚢胞腎
- 腎臓に、液体が詰まった袋状の構造(嚢胞:のうほう)が多数発生し、腎臓がふくれあがる疾患。多くの場合、腎不全に至る。遺伝病であり、尿細管の一次繊毛に発現するTRPチャネルの遺伝子の異常が原因である。
- 注6) フィードバックシステム
- 自分の活動の結果をもとにして、次の自分の活動を制御する方式のこと。
- 注7) 気管繊毛上皮
- 気管の内側の表面にある繊毛細胞が集まってできた上皮組織のこと。気管に入ってきた異物を繊毛の表面にある粘膜に付着させ、繊毛運動によって排除する。
- 注8) RNA干渉
- 遺伝子が発現する時に生成されるメッセンジャーRNAを切断することにより、その遺伝子の発現を抑制する実験手法をいう。
- 注9) 受容器電位
- 生物の感覚器では、刺激を受けるとその受容体が細胞の膜電位の変化を引き起こす。この電位を受容器電位という。
- 注10) 電位依存性カルシウムチャネル
- 鞭毛・繊毛の運動をはじめ、多くの生物現象はカルシウムイオンによって制御されている。細胞外からのカルシウムイオンの流入の経路となる膜たんぱく質がカルシウムチャネルと呼ばれ、その中でも、細胞膜の電位を感じて開くタイプを電位依存性カルシウムチャネルという。本プロジェクトの研究により、クラミドモナスでの分子実体がCAV2というたんぱく質であり、鞭毛の根元を避ける分布を示すことが明らかになっている。
<論文名>
“Mechanoreception in Motile Flagella of Chlamydomonas”
(クラミドモナスの運動能のある鞭毛による機械受容)
doi: 10.1038/ncb2214
Kenta Fujiu, Yoshitaka Nakayama, Hidetoshi Iida, Masahiro Sokabe, and Kenjiro Yoshimura
<お問い合わせ先>
<研究に関すること>
曽我部 正博(ソカベ マサヒロ)
名古屋大学 大学院医学系研究科 細胞生物物理学 教授
JST 戦略的創造研究推進事業 発展研究(ICORP-SORST)
「ナノ・マイクロ超分子複合体によるメカノトランスダクション機構の解明プロジェクト」 総括
Tel:052-744-2051
E-mail:
<JSTの事業に関すること>
金子 博之(カネコ ヒロユキ)
科学技術振興機構 イノベーション推進本部 研究プロジェクト推進部
〒102-0075 東京都千代田区三番町5 三番町ビル
Tel:03-3512-3528 Fax:03-3222-2068
E-mail: