ポイント
- ほ乳類の大脳皮質が作られる際に神経細胞は、「ロコモーション様式」と呼ばれる特徴的な移動様式で特定の位置まで移動し、大脳皮質の6層構造を形成する。
- 神経細胞の増殖を停止させる分子Cdk5、p27と、脳疾患の原因遺伝子Dcxとが「ロコモーション様式の移動」を制御していることが分かった。
- 神経細胞の「増殖しない」という特徴を決定づける分子が、増殖停止後に新たな機能を獲得し、これが脳の高度な組織化過程に重要であることが示された。
JST 戦略的創造研究推進事業において、JSTの川内 健史 さきがけ研究者(兼・慶應義塾大学 医学部・特任講師)らは、ほ乳類の大脳皮質が作られる際、神経細胞が他の細胞には見られない特殊な様式(ロコモーション様式)で移動する仕組みの一端を明らかにしました。
大脳皮質が作られる際、神経前駆細胞注1)から分化した神経細胞は、細胞分裂を停止した後、(1)神経突起の根元に「特徴的な膨らみ」を形成し、さらに、(2)細胞核が細長く伸びて「膨らみ」の中に入り込む過程を繰り返すロコモーション様式の移動を行います。このような神経細胞の移動は、ほ乳類に特異的な6層構造の大脳皮質が作られるために重要な発生段階であり、これが破綻すると、てんかんや知的障害を伴う滑脳症注2)などのさまざまな脳疾患を引き起こします。
川内研究者は、神経細胞の増殖を停止させる分子p27、滑脳症の原因遺伝子Dcx、これらの上流制御因子Cdk5の機能を抑えると、いずれの場合もロコモーション様式の(1)と(2)の両方の段階が阻害され、神経細胞が正しく移動できなくなることを明らかにしました。
以前の研究から、Cdk5とp27は、神経細胞の増殖停止や複雑な形態変化に関与すると分かっていました。今回の結果から、Cdk5とp27が、神経細胞の特徴を規定するだけでなく、高度な脳の組織化を促す一連のステップすべてを制御する司令塔のような分子基盤であることが示されました。
本研究成果は、2014年9月2日(英国時間)に発生生物学専門誌「Development」のオンライン版で公開されます。
本成果は、以下の事業・研究領域・研究課題によって得られました。
戦略的創造研究推進事業
| 研究領域 |
「脳神経回路の形成・動作と制御」
(研究総括:村上 富士夫 大阪大学 大学院生命機能研究科 特任教授) |
| 研究課題名 |
「細胞内機能ドメインが大脳皮質形成に果たす役割の解明」 |
| 研究者 |
川内 健史(JSTさきがけ研究者) |
| 研究実施場所 |
慶應義塾大学 医学部 |
| 研究期間 |
平成21年10月~平成27年3月 |
<研究の背景と経緯>
記憶や学習といった生物の行動は、脳の働きによって制御されています。脳は、機能に応じて厳密な組織化がなされており、高次脳機能を担う大脳皮質の場合、ほ乳類では6つの層に分かれています(図1)。このような6層構造の大脳皮質が形成されるためには、(1)神経前駆細胞(神経幹細胞)が細胞分裂を止めて、神経細胞へと分化する、(2)誕生した神経細胞が長い距離を移動して適切な場所へ配置されることが必要です(図1,図2)。そして、これらの発生過程に異常をきたすと脳が正しく機能できなくなってしまいます。例えば、(2)の神経細胞移動が障害されると、滑脳症(てんかんや知的障害を伴う脳奇形)などのさまざまな脳疾患が引き起こされてしまうと考えられています。
細胞の移動は、脳に限らず他の多くの組織の発生過程や、がんなどの疾患においても観察されますが、その移動様式は多種多様であることが知られています(図3)。その中でも、発生期の大脳皮質の神経細胞は、他の細胞では見られない特殊な移動様式である「ロコモーション様式の移動」を示します。具体的には、(1)進行方向前方の突起の根元に「特徴的な膨らみ」が形成され、(2)細胞核が細長く伸びて、その「膨らみ」の中へと入り込んでいく、という過程を繰り返しながら、神経細胞は最終的に配置される場所まで動いていき、その結果、秩序だった6層構造の大脳皮質が形成されます(図3A,図4)。
突起の根元に形成される「特徴的な膨らみ」は、移動している神経細胞のみに観察され、静止している神経細胞や、神経細胞以外の移動細胞ではみられない構造体です。このことから、神経細胞に特異的に発現する分子が、この「膨らみ」の形成を制御していることは予想されましたが、神経細胞に強く発現する分子は数多く知られているため、どの分子が「突起の膨らみ」の形成を制御しているのかについては、分かっていませんでした。
<研究の内容>
大脳皮質形成において、神経細胞はロコモーション様式の移動を行う前に、複雑な形態変化を起こします(図1,図2)。このため、ロコモーション様式の移動(図2の(2)の段階)を直接解析することは困難でした。最近、川内研究者らはこの複雑な形態変化の影響を排して、ロコモーション様式の移動を直接解析できる新たな実験系を開発しました(参考文献1)。これは、マウス大脳皮質の組織スライスを顕微鏡下で培養し、組織スライスの中をロコモーション様式で移動する神経細胞を観察しながら、適当なタイミングで特定の分子に対する阻害剤を添加し、引き続き同じ神経細胞の挙動を観察することで、どのような影響がでるかなどを調べることができます。そこでこの独自の実験系などを用いて、神経細胞がロコモーション様式の移動の際に形成する「特徴的な膨らみ」(図3Aの(1)の段階)や「核の伸長」(図3Aの(2)の段階)を制御する分子を探索しました。その結果、神経細胞で強く活性化しているキナーゼ(リン酸化酵素)であるCdk5に対する阻害剤を添加すると、ロコモーション様式で移動する神経細胞の「特徴的な膨らみ」の形成と「核の伸長」が、どちらも抑制され、結果として神経細胞が正しく移動できなくなることが分かりました(図5)。以前に川内研究者らは、Cdk5キナーゼの基質の1つとして、神経前駆細胞が増殖を停止して神経細胞へと分化する過程(図2の(1)の段階)を制御している分子p27を同定していたことから(参考文献2)、次に遺伝子ノックダウン法によりp27の発現量を減らしたところ、Cdk5の阻害剤を添加した場合と同様に、ロコモーション様式で移動する細胞の「特徴的な膨らみ」の形成と「核の伸長」が抑制されました。これらより、神経細胞が増殖を停止するために必要な分子であるp27と、その上流制御因子であるCdk5が、神経細胞に特徴的な移動様式である「ロコモーション様式の移動」(図2の(2)の段階)を制御していることが分かりました(図5,図7)。
Cdk5は、p27に加えて、Dcxという分子もリン酸化し、Dcxの活性を負に制御することも知られています。そこで川内研究者らは、Dcxについても同様の実験系を用いて解析を行いました。Dcxを過剰に発現して機能を亢進する実験と、遺伝子ノックダウン法によりDcxの発現量を減らす実験を行ったところ、どちらの場合も神経細胞の移動速度が低下しました。さらに、Dcxの発現量を増やした場合も減らした場合も、ロコモーション様式で移動中の神経細胞の「特徴的な膨らみの形成」(図3Aの(1)の段階)と「核の伸長」(図3Aの(2)の段階)が抑制されました。これらより、Dcxが適切な活性を保つことが、ロコモーション様式の移動に重要であることが分かりました(図6,図7)。
以上のことから、神経前駆細胞の増殖を停止させ、神経細胞を生み出すために必要な分子であるCdk5、p27が、Dcxと協調的に働くことにより、ロコモーション様式の移動における「特徴的な膨らみの形成」(図3Aの(1)の段階)と「核の伸長」(図3Aの(2)の段階)の両方を制御していることが明らかとなりました(図7)。
<今後の展開>
本研究により、Cdk5とp27が、脳が作られるために重要なステップである、神経細胞の増殖停止、複雑な形態変化、ロコモーション様式の移動という一連の流れをすべて制御する司令塔のような役割を果たすことが分かりました。
p27は、神経細胞に限らず多くの細胞の増殖停止を制御する鍵分子であるにも関わらず、なぜ神経細胞において特別の機能を獲得するのかについてはまだ明確な答えはありませんが、神経細胞で強く活性化するCdk5と同時に働くことが、重要である可能性が考えられます。
さらに本研究では、Cdk5とp27が、Dcxと協調して働くという興味深い結果も得られました。Dcxは、滑脳症の原因遺伝子としても知られています。今回、Dcxの役割が解明されたことにより、滑脳症をはじめとした脳疾患の原因究明が進む可能性もあります。
私たちの脳の中にある、ほとんどすべての神経細胞は、発生期に産生されます。そして、分化した神経細胞は、一生涯、増殖することはありません。すなわち、老化や何らかの傷害によって神経細胞が失われたとしても、神経細胞を補うことはできません。なぜ、このようなリスクの高い特徴をもっているのかについては、未解明な点が多く残されています。近年、大人の脳で、神経細胞が増殖を再開してしまうことは、アルツハイマー病などの神経変性疾患の引き金になるという仮説が提唱されています。すなわち、増殖を停止することは、神経細胞が生存するためにも必要である可能性が考えられます。「増殖を停止させる分子」が、神経細胞の成熟や機能維持においても重要な役割を果たしていることと関係があるかもしれません。将来、本研究の成果が、神経細胞が積極的に増殖を停止する理由の解明につながることが期待されます。
<付記>
本研究は、同志社大学 脳科学研究科の西村 嘉晃 助教、慶應義塾大学 医学部の鹿内 弥磨 特任研究員、慶應義塾大学 医学部の仲嶋 一範 教授、愛知県心身障害者コロニー発達障害研究所の永田 浩一 研究部長、国立精神・神経医療研究センター 神経研究所の星野 幹雄 研究部長、先端医療研究財団の鍋島 陽一 先端医療センター長、同志社大学 脳科学研究科の水谷 健一 准教授、早稲田大学 先進医工学部の大島 登志男 教授との共同で行ったものです。
<参考図>
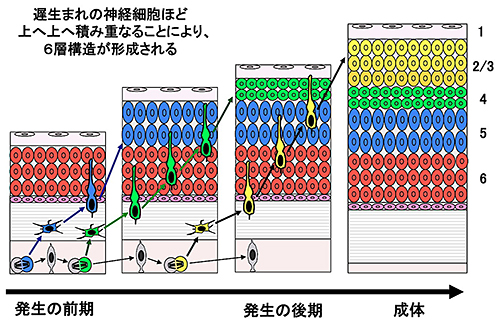
図1 ほ乳類の大脳皮質6層構造の形成過程
ほ乳類の大脳皮質は、6層構造を成す(一番右の図)。この層構造が形成されるには、適切なタイミングで増殖を停止した神経細胞が脳の表層へ向かって移動し、正しく配置されることが重要である。つまり、脳の内側(脳室側:図の下側)で誕生した遅生まれの細胞(黄色の細胞)が、すでに移動を終了した早生まれの細胞(青色の細胞と緑色の細胞)を追い越し、さらに上に積み重なっていくことにより、6層構造が形成される。なお、マウスは2層と3層の区別がないため、一般に「2/3層」と表記する。
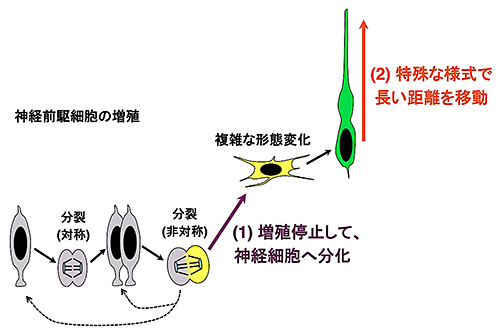
図2 神経細胞は増殖を停止した後、長い距離を移動する
神経前駆細胞(灰色の細胞)は活発に分裂するが、神経細胞へ分化すると同時に、増殖を停止する(図の(1))。増殖を停止した神経細胞は、複雑な形態変化を示した後(黄色の細胞)、他の細胞には見られない特殊な様式(図3Aを参照)で、脳の表層までの長い距離を移動する(緑色の細胞)(図の(2))。
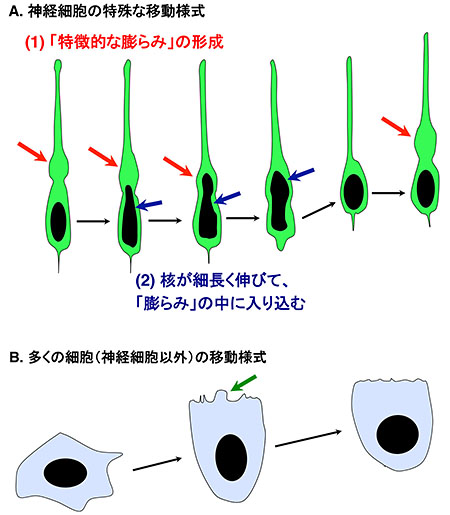
図3 神経細胞に特徴的な「ロコモーション様式の移動」
- (A) 神経細胞は、移動過程の大半において、図3Aに示すような「ロコモーション様式」呼ばれる移動様式をとる。(1)まず、太い神経突起の根元に「特徴的な膨らみ」(赤色の矢印)を形成する。(2)その後、図中に黒色で示した細胞核が細長く伸びていき(青色の矢印)、伸びた核は「膨らみ」の中へと入り込んでいく。このような形態変化を繰り返しながら、神経細胞は脳の表層へ向かって移動する。
- (B) 一般的な多くの細胞の移動様式。まず、細胞の膜が前方に突出し(緑色の矢印)、その後、細胞核が前方へと移動する。神経細胞以外の細胞は、移動過程で、「特徴的な膨らみ」を形成することはない。
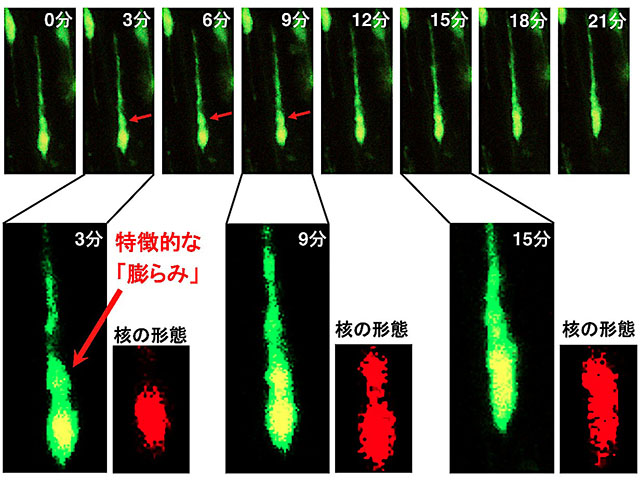
図4 培養した大脳皮質組織スライス内を移動する神経細胞の経時観察
培養した大脳皮質組織のスライス内を移動する神経細胞(緑色)を、3分おきに観察した。観察の3分後から、突起の根元に「膨らみ」が形成され(赤色の矢印)、その後、細胞核が細長く伸びていく(細胞核:黄色。ただし、下段の写真で「核の形態」と表記した拡大写真では赤色)。
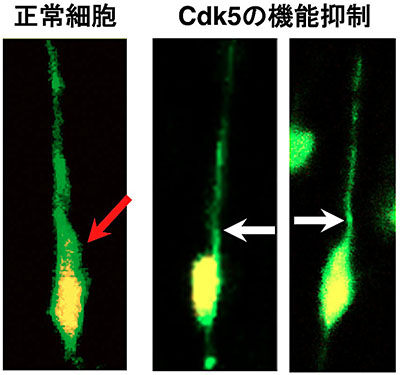
図5 Cdk5の機能抑制による
「膨らみ」の形成の阻害
培養した大脳皮質組織のスライス内を移動する正常な神経細胞(左の細胞)と、Cdk5の阻害剤を添加した組織スライス内の神経細胞(中央と右の細胞)。正常細胞では、突起の根元の「膨らみ」が観察されるが(赤色の矢印)、Cdk5の機能を阻害すると、突起が細くなり、根元の「膨らみ」もなくなる(白色の矢印)。緑色が神経細胞、黄色はその細胞核。
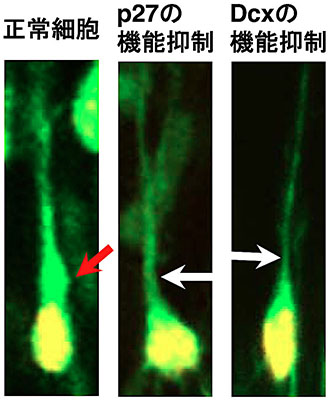
図6 p27もしくはDcxの
機能抑制による「膨らみ」の形成阻害
培養した大脳皮質組織のスライス内を移動する正常な神経細胞(左の細胞)と、p27の機能を阻害した神経細胞(中央の細胞)と、Dcxの機能を阻害した神経細胞(右の細胞)。正常細胞では、突起の根元の「膨らみ」が観察されるが(赤色の矢印)、p27もしくはDcxの機能を阻害すると、突起が細くなり、根元の「膨らみ」もなくなる(白色の矢印)。緑色が神経細胞、黄色はその細胞核。
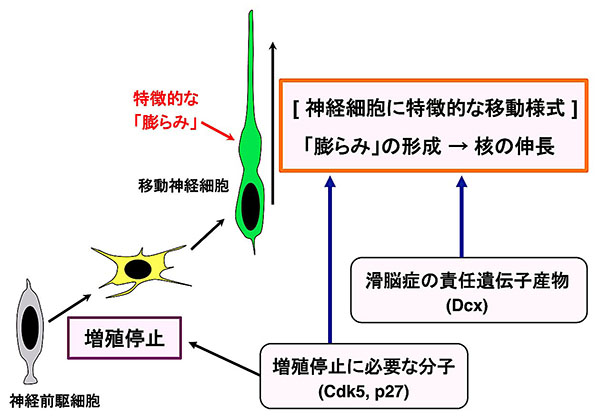
図7 本研究の概念図
神経細胞の増殖を停止させるCdk5、p27が、ロコモーション様式で移動する神経細胞に観察される「特徴的な膨らみ」の形成や、細胞核が細長く伸びる過程を制御することにより、ロコモーション様式の移動にも関与していることが分かりました。また、これらの分子は、滑脳症の原因遺伝子Dcxと協調的に働いていることも明らかになりました。
<用語解説>
- 注1) 神経前駆細胞
- 主に発生期の脳に存在し、神経細胞を産生する細胞。神経前駆細胞は、活発に細胞分裂して自己増殖するとともに、主に非対称分裂により、神経細胞を産生する(図2の灰色の細胞を参照)。神経前駆細胞から産生された神経細胞は、増殖を停止して、一生涯、再び分裂することはない。
- 注2) 滑脳症
- 脳の一部もしくは全体で、「脳のしわ」がなくなる脳奇形(疾患名は、脳の表面が滑らかになることに由来する)。脳のごく一部のみでしわがなくなる軽微なものもあるが、多くの場合は、てんかんや知的障害を伴う。ヒトおよびモデルマウスなどを用いた解析から、主に神経細胞移動が障害されたことが原因と考えられている。
- なお、Dcxの変異は、男性ではX染色体連鎖型の滑脳症を引き起こすが、女性では、同じように神経細胞移動の異常が原因である「皮質下帯状異所性灰白質」という病気になる。
<論文タイトル>
“Cdk5 and its substrates, Dcx and p27kip1, regulate cytoplasmic dilation formation and nuclear elongation in migrating neurons”
(Cdk5とその基質であるDcxとp27kip1は、移動神経細胞における細胞質の「膨らみ」の形成と核の伸長を制御する)
doi: 10.1242/dev.111294
<参考文献>
- 参考文献1:
Nishimura YV et al. “Dissecting the factors involved in the locomotion mode of neuronal migration in the developing cerebral cortex”
J. Biol. Chem. 285, 5878-5887 (2010).
- 参考文献2:
Kawauchi T et al. “Cdk5 phosphorylates and stabilizes p27kip1 contributing to actin organization and cortical neuronal migration”
Nature Cell Biol. 8. 17-26 (2006).
<お問い合わせ先>
<研究に関すること>
川内 健史(カワウチ タケシ)
科学技術振興機構 さきがけ研究者
慶應義塾大学 医学部生理学教室 特任講師
〒160-8582 東京都新宿区信濃町35
Tel:03-5363-3743 Fax:03-5379-1977
E-mail:
<JST事業に関すること>
松尾 浩司(マツオ コウジ)、川口 貴史(カワグチ タカフミ) 、眞後 俊幸(シンゴ トシユキ)
科学技術振興機構 戦略研究推進部 ライフイノベーショングループ
〒102-0076 東京都千代田区五番町7 K’s五番町
Tel:03-3512-3525 Fax:03-3222-2067
E-mail:
<報道担当>
科学技術振興機構 広報課
〒102-8666 東京都千代田区四番町5番地3
Tel:03-5214-8404 Fax:03-5214-8432
E-mail: