奈良先端科学技術大学院大学(学長:横矢 直和) 先端科学技術研究科 バイオサイエンス領域の伊藤 寿朗 教授、山口 暢俊 助教(兼任:JST 戦略的創造研究推進事業 さきがけ研究者)、立命館大学 立命館グローバル・イノベーション研究機構 菅野 茂夫 助教(兼任:JST 戦略的創造研究推進事業 さきがけ研究者)、東北大学 大学院生命科学研究科の西谷 和彦 教授、名古屋大学 大学院生命農学研究科の榊原 均 教授らの共同研究グループは、花の中心部にあり、受粉して果実や種子になる「めしべ注1)」が形成されるときに、その形成に関わる遺伝子を働かせるスイッチ(遺伝子発現)がオンになる詳細な仕組みを世界に先駆けて明らかにしました。この成果により、人工的にめしべの大きさや数などが調節できるようになれば、環境に応じた食糧の増産や安定供給などが期待できます。
めしべをつくるためには、遺伝子発現のスイッチをオンにする複数の転写因子注2)(タンパク質)が働くことは分かっていました。このスイッチが入っていない状態では、遺伝子の本体である長いDNA注3)はタンパク質であるヒストン注4)に巻きつき、クロマチン注5)と呼ばれるDNAが折りたたまれて閉じた構造を作ります。このため、転写因子は目標のDNAにたどり着けず、めしべがつくられません。遺伝子発現のスイッチをオンにするためには、その構造をほどき、開いていく必要があります。しかしながら、「それらの転写因子がどのような順序や方法でクロマチンに働きかけ、その構造と遺伝子の発現を変化させていくのか?」について詳しい仕組みについては謎でした。
伊藤教授らの共同研究グループは、モデル植物のシロイヌナズナ注6)を使って実験を重ねた結果、最初の段階で転写因子(パイオニア転写因子)が、クロマチンの構造を変化させる因子(クロマチンリモデリング因子)と共に働いて、複雑な構造をほどくことを見いだしました。次いで別の転写因子がめしべ形成のDNAに直接結合して発現させるという、2つの転写因子が連係する順序や仕組みを突き止めました。このような仕組みはこれまでは動物でのみ報告されており、今回、植物にも同様の仕組みが存在し、めしべをつくるために必要であると解明したことは、植物の進化や生き残り戦略を知る上でも重要です。
本研究の成果は2018年12月11日付けで「Nature Communications」オンライン版に掲載されます。
本研究は日本学術振興会(JSPS) 科学研究費補助金、科学技術振興機構(JST) 戦略的創造研究推進事業 個人型研究(さきがけ)の支援を受けて行われました。
<背景>
植物は生殖器官であるめしべをつくって、次の世代の子孫を残します。めしべをつくるためには、遺伝子発現のスイッチをオンにする転写因子が働きます。この転写因子は遺伝子発現のオン、オフを切り替えて、形づくりを進めます。遺伝子発現のスイッチが入っていないオフの状態では、長いDNAはタンパク質であるヒストンに巻きついて、クロマチンと呼ばれるDNAが折りたたまれた構造を作っています。遺伝子発現のスイッチをオンにするためには、その構造をほどき、開いていく必要があります。これまでにめしべをつくるために関わる転写因子が複数分かっていましたが、それらの転写因子がどのような順序や方法でクロマチンに働きかけて、その構造と遺伝子の発現を変化させていくのかについて、詳しい仕組みは未解明でした。
<研究手法と成果>
はじめに本研究グループは、モデル植物であるシロイヌナズナを用い、めしべをつくる機能がある転写因子AGAMOUS(AGタンパク質注7))とCRABS CLAW(CRCタンパク質注8))に注目しました。AGタンパク質が作用しないag突然変異体では、めしべはできなくなることが分かっていたからです。また、CRCタンパク質が働かなくなったcrc突然変異体でも、正常なめしべはできないことが報告されていました。そこで、2つの突然変異体を用いて、遺伝子の発現を網羅的に解析したところ、AGタンパク質とCRCタンパク質のそれぞれが複数の遺伝子の発現を同じように調節することを発見しました。それらで調節される遺伝子の中に、植物ホルモン注9)の1つであるオーキシン注10)を合成する働きがあるYUCCA4(YUC4)遺伝子注11)が含まれていました。
オーキシンは形づくりを実行する機能があるため、AGタンパク質とCRCタンパク質のYUC4遺伝子への発現の調節について調べたところ、AGタンパク質とCRCタンパク質がつくれないagとcrc突然変異体ではYUC4遺伝子の発現量は減少することが分かりました。逆に、AGタンパク質やCRCタンパク質の活性を強くした場合には、YUC4遺伝子の発現量は増加しました。また、AGタンパク質、CRCタンパク質のどちらもYUC4遺伝子に直接結合することも発見しました。この結果から、AGタンパク質とCRCタンパク質はどちらもYUC4遺伝子に結合して発現のスイッチを入れることを明らかにしました。
しかし、AGタンパク質とCRCタンパク質はどちらもYUC4遺伝子に結合していたものの、両者の結合部位は少しずれていました。またAGタンパク質とCRCタンパク質の蓄積部位も完全には一致しませんでした。そのため、AGタンパク質とCRCタンパク質は1つの複合体を形成して同時にYUC4遺伝子の同じ位置付近に結合するのではなく、結合する順序があるのではないかと予想しました。AGタンパク質はクロマチンの構造を変化させる働きがあるクロマチンリモデリング因子CHROMATIN-REMODELING PROTEIN 11(CHR11)やCHR17と複合体をつくるというこれまでの報告に注目して調べたところ、さらにCHRタンパク質注12)も、AGタンパク質やCRCタンパク質と同じようにYUC4遺伝子に直接結合して、発現のスイッチを入れるように作用することを発見しました。CHRタンパク質は動植物を問わず存在するタンパク質であることが知られており、動物の相同タンパク質(同じ祖先遺伝子に基づく類似したタンパク質)ではDNAに巻きついているヒストンを移動させ、クロマチンの構造を開くように働きます。そこで本研究グループは、クロマチンの構造を解析する実験手法「FAIRE注13)」を用いて、AGタンパク質とCHRタンパク質がYUC4遺伝子を含むクロマチンの構造をほどくことを明らかにしました。さらに、AG、CHR、CRCそれぞれのタンパク質がクロマチンに作用する順番を調べたところ、AGタンパク質とCHRタンパク質の複合体がはじめにクロマチンに作用してそのDNAの構造を開き、CRCタンパク質をDNAにアクセス可能にする順序を明確にすることに成功しました(図1)。
このようなクロマチン構造の変化によって、めしべができる部位でYUC4遺伝子が発現を開始します。そのあと、オーキシンの合成、幹細胞の分裂停止、細胞壁注14)の構築が行われ、めしべの形づくりが実行されていきます(図2)。
<波及効果>
パイオニアとなる転写因子がクロマチンリモデリング因子と共に働いてクロマチンの構造を変え、次の転写因子が機能できるようにする仕組みは動物では見つかっていましたが、今回、本研究グループは植物にも同様の仕組みが存在し、めしべをつくるために必要であることを世界に先駆けて発見しました。また、複数の転写因子の働きを変化させることで、めしべからつくられる果実や種子の大きさや数などさまざまな要因を最適に人為調節できる可能性があり、食糧増産や安定供給が期待されます。
<参考図>
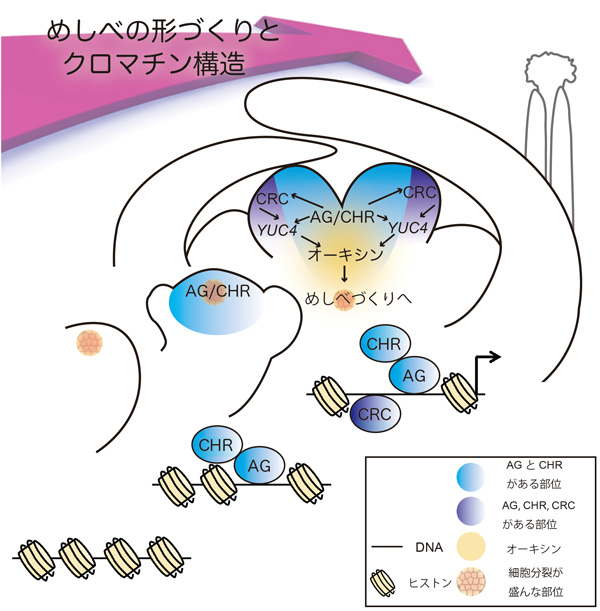
図1 めしべの形づくりとクロマチン構造
めしべの形づくりが進むにつれて、転写因子の働きによってYUC4遺伝子のクロマチンの構造が開く。するとめしべがつくられる。
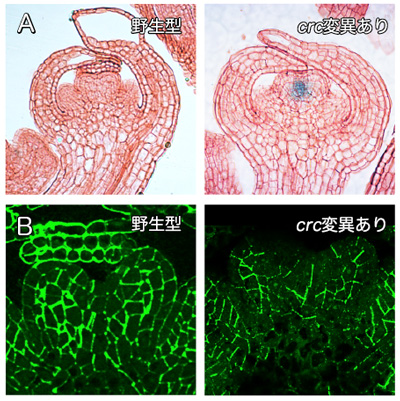
図2 野生型とcrc変異を持つめしべ原基での細胞分裂と細胞壁の様子
分裂停止が起こっている野生型と分裂停止が起こっていないcrc変異体を用いて、細胞分裂と細胞壁の観察を行った。
- (A)細胞分裂を観察した写真。分裂活性が高い部分が青くなっている。Aの実験結果から、crc変異体では細胞分裂が盛んな様子が分かる。
- (B)細胞壁を観察した写真。細胞を取り囲む細胞壁が緑色になっている。Bの実験結果から、crc変異体では細胞壁の構築が適切に行われていない様子が分かる。
<用語解説>
- 注1)めしべ
- 被子植物の花にある雌性の生殖器官。受粉して実や種子になる。
- 注2)転写因子
- 遺伝子の発現のオン、オフを切り替えるタンパク質。DNA上の特定の塩基配列に結合し、遺伝子の発現量を調節する。
- 注3)DNA
- 細胞の核内で長い鎖状の2重らせん構造を形成し、生物の遺伝情報を保持している物質。
- 注4)ヒストン
- 長いDNAを折りたたんで核内に収納するタンパク質。
- 注5)クロマチン
- DNAをヒストンに巻きコンパクトに核内に収納する構造。遺伝子発現制御においても重要な役割を持っている。
- 注6)シロイヌナズナ
- 遺伝子の解析を行うのに適したアブラナ科の1年草。
- 注7)AGタンパク質
- 花の器官分化などに関わり、MADSボックスと呼ばれるDNA結合領域を持つ転写因子。
- 注8)CRCタンパク質
- めしべの形成や、花の幹細胞の増殖抑制などに関わり、コードする遺伝子にYABBYドメイン領域を持つ転写因子。
- 注9)植物ホルモン
- 植物の体内で作られ、成長を調節する低分子量有機化合物。
- 注10)オーキシン
- 器官の形成を促進する働きがある低分子量有機化合物。
- 注11)YUC4遺伝子
- オーキシンを合成する酵素をコードする遺伝子。
- 注12)CHRタンパク質
- DNAに巻きついているヒストンを移動させ、クロマチンの構造を開くように働くクロマチンリモデリング因子。
- 注13)FAIRE
- クロマチンの構造を調べる実験手法。増幅したDNAの量が多い場合には、クロマチンが開いていることを意味する。
- 注14)細胞壁
- 細胞膜の外側に位置する構造。細胞の改築、補強、防御など多くの役割を果たす。
<掲載情報>
| タイトル |
“Chromatin-mediated feed-forward auin biosynthesis in floral meristem determinacy” |
| 著者 |
Nobutoshi Yamaguchi, Jiangbo Huang, Yoshitaka Tatsumi, Masato Abe, Shigeo S. Sugano, Mikiko Kojima, Yumiko Takebayashi, Takatoshi Kiba, Ryusuke Yokoyama, Kazuhiko Nishitani, Hitoshi Sakakibara, and Toshiro Ito*
*責任著者
|
| DOI |
10.1038/s41467-018-07763-0 |
<お問い合わせ先>
<研究に関すること>
伊藤 寿朗(イトウ トシロウ)
奈良先端科学技術大学院大学 先端科学技術研究科 バイオサイエンス領域 花発生分子遺伝学研究室 教授
Tel:0743-72-5500 Fax:0743-72-5502
E-mail:
研究室紹介ホームページ:http://bsw3.naist.jp/courses/courses112.html
研究室ホームページ:http://bsw3.naist.jp/ito/
<JST事業に関すること>
川口 哲(カワグチ テツ)
科学技術振興機構 戦略研究推進部
Tel:03-3512-3525 Fax:03-3222-2064
E-mail:
<報道担当>
奈良先端科学技術大学院大学 企画総務課 広報渉外係
Tel:0743-72-5026 Fax:0743-72-5011
E-mail:
科学技術振興機構 広報課
Tel:03-5214-8404 Fax:03-5214-8432
E-mail: