ポイント
- 知識は複数の記憶情報を共有化することで形成されるが、個々の記憶のアイデンティティがどのような仕組みで保たれているのかは不明だった。
- マウスを用いて、共有化された記憶を担う細胞(エングラム細胞)が持つ異なるシナプスの伝達効率の制御が、それぞれの記憶のアイデンティティを担うことを明らかにした。
- 記憶のアイデンティティの喪失は、記憶障害を伴う認知症などに密接に関わっていることから、本成果はこれらの疾患の予防・治療法の創出にもつながると期待される。
JST 戦略的創造研究推進事業において、富山大学 大学院医学薬学研究部(医学) 生化学講座の井ノ口 馨 教授らは、マウスで、関連付けられた記憶それぞれのアイデンティティ注1)は、シナプス特異性が担っていることを世界で初めて明らかにしました。
記憶は特定の神経細胞集団として脳内に蓄えられています。記憶同士が相互作用するときには、それぞれの記憶に対応する記憶エングラム細胞注2)が共有化され、その結果として元々独立した記憶同士が関連付けられ新しい意味を持つ記憶(知識)が形成されます。一方で、これまでの研究において、それぞれの記憶が相互作用しても、元々の記憶のアイデンティティは保たれていますが、どのような仕組みによりアイデンティティが保たれているのかは不明のままでした。
本研究グループは、まず、異なる音恐怖記憶を用いて、2つの記憶の相互作用は扁桃体注3)に存在するエングラム細胞の共有化が担っていることを示しました。一方で、それぞれの記憶のアイデンティティは、聴覚皮質と扁桃体の間に形成される特異的なシナプス可塑性注4)が担っていることを示しました。すなわち、2つの記憶を持つ扁桃体のエングラム細胞上に存在する異なるシナプスが異なる記憶を担っていることを明らかにしました。また、この研究の過程で、完全な逆行性健忘注5)はエングラムの消失を伴い記憶そのものが消え去っていることも明らかになりました。
記憶がアイデンティティを保つ仕組みに関する今回の研究は、一つ一つの記憶を正確に保存しつつ記憶同士を関連付けて知識や概念を形成していくヒトの高次脳機能の解明につながる成果です。また、記憶のアイデンティティの喪失は、記憶障害を伴う認知症などに密接に関わっていることから、本成果はこれらの疾患の予防法や治療法の創出につながると期待されます。
本研究は、富山大学 大学院医学薬学研究部(医学)のカリム・アブドウ 特命助教と自治医科大学の村松 慎一 教授らと共同で行ったものです。
本研究成果は、2018年6月15日(米国東部時間)に米国科学誌「Science」のオンライン版で公開されます。
本成果は、以下の事業・研究領域・研究課題によって得られました。
戦略的創造研究推進事業 チーム型研究(CREST)
| 研究領域 |
「生命動態の理解と制御のための基盤技術の創出」
(研究総括:山本 雅 沖縄科学技術大学院大学 教授) |
| 研究課題名 |
「細胞集団の活動動態解析と回路モデルに基づいた記憶統合プロセスの解明」 |
| 研究代表者 |
井ノ口 馨(富山大学 大学院医学薬学研究部(医学) 教授) |
| 研究期間 |
平成25年10月~平成31年3月 |
上記研究課題では、記憶のダイナミックな側面に注目し、ニューロン集団の活動動態に焦点を当てた解析と数理モデルの構築を通して、神経回路レベルの記憶のメカニズム解明を目指します。
<研究の背景と経緯>
異なる経験により形成される複数の記憶情報を関連する記憶として連合させ、新しい意味を持った記憶(知識)として貯蔵することは、知識や概念の形成という観点から注目を集めています。
近年、個々の記憶は経験時に活動した特定のエングラム細胞集団として脳内に蓄えられていること、また、記憶同士が相互作用するときには、それぞれの記憶をつかさどるエングラム細胞を両方の記憶が共有し、その結果として記憶同士の間に関連付けが形成されることが明らかになってきました。一方で、それぞれの記憶が相互作用しても、元々の記憶のアイデンティティは保たれていますが、どのような仕組みによりアイデンティティが保たれているのかは不明のままでした。
<研究の内容>
他の記憶と相互作用しつつ個々の記憶のアイデンティティを保持するメカニズムの解明に「シナプス特異性」の観点から取り組みました。
逆行性健忘を引き起こす実験系の構築
本研究では、マウスを用い、まず、再固定化の阻害注6)により、音恐怖記憶の逆行性健忘を引き起こす実験系を構築しました。
条件刺激注7)として7kHzの音、無条件刺激注7)として電気ショックを組み合わせて条件付けを行い、1日後に7kHzの音の提示で音恐怖記憶を想起させた(テスト1)直後に、この記憶の責任領域の扁桃体に種々の薬液を注入し、その効果を測定しました(図1A)。記憶の想起直後にアニソマイシン注8)を投与し、たんぱく質合成を阻害した群では、次の日に音提示により恐怖反応であるフリージング(すくみ反応)が低下しており(テスト2)、部分的な健忘が引き起こされました。想起に伴う記憶の減弱化を促進することが知られているタットベクリン注9)をアニソマイシンと共に投与した群では、ほとんどすくみ反応を示さず、完全な健忘が引き起こされました(図1B)。
また、条件付けの時に活動した扁桃体の神経細胞をoChIEF注10)で標識しておくことで(図1A左端)光照射のみで扁桃体のエングラム細胞を人為的に活性化することができるようにしました。そこで、逆行性健忘を引き起こした翌日に、音刺激のない状態で光照射をすると(図1、テスト3)、アニソマイシン投与群では健忘を引き起こしていない実験対照群と同程度の高いすくみ反応を示しました。一方でタットベクリン+アニソマイシン投与群では、光照射しても全くすくみ反応を示しませんでした(図1C)。これらの結果より、部分的な健忘ではエングラムは残っているのに対して、完全な健忘状態ではエングラムは消失しており、記憶そのものが消え去っていることが強く示唆されました。
記憶のアイデンティティを保つ仕組み
マウスに、扁桃体を必要とする学習である2種類の音による恐怖条件付けとして、最初に「7kHzの音+電気ショック」、次にその5時間後または24時間後に「2kHzの音+電気ショック」を行いました。学習後、マウスは7kHz、2kHzのいずれの音を聞いても恐怖反応であるフリージング(すくみ反応)を示すようになりました(図2A)。2種類の条件付けを与える間隔が、実験対照群として用いた24時間の場合はどちらの音に対しても同程度のすくみ反応を示しましたが、実験群の5時間の場合は後で条件付けに用いた2kHzのブザー音に対するすくみ反応が増大しました。この結果は、短い間隔の条件付けでは、2つの記憶の間に相互作用が形成されたことを示しています。
その際に、扁桃体領域において、各記憶に応答して活動したエングラム細胞集団をcatFISH法注11)により特定したところ、5時間間隔で2つの条件付けを行った群では両記憶を担うエングラム細胞集団が60パーセントという非常に高い共有率を示し(図2B)、共有化されたエングラム細胞集団が両記憶の関連付けを担っていることを示しています。
その一方で、7kHzの音で条件付けされたマウスは2kHzの音に対して、2kHzのブザー音で条件付けされたマウスは7kHzの音に対しては、いずれもすくみ反応を示さなかったことから(図2C)、それぞれの記憶は区別されており、記憶のアイデンティティは保たれていることが分かりました。
これに対して音情報をつかさどる大脳皮質の聴覚野においては、それぞれの音に応答して活動した音のエングラム細胞集団は低い共有率(20パーセント)を示し、2つのエングラム細胞群が独立していることが分かりました(図2B)。
以上から、扁桃体では2つの音恐怖記憶が同じエングラム細胞群により担われているのに対して、聴覚野では2つの音記憶は異なるエングラム細胞群が担っていることが分かりました(図2D)。これらの結果から、聴覚野の2種類のエングラム細胞群は、扁桃体の同一の音恐怖エングラム細胞上にシナプスを形成していることが想定されました。
記憶の同一性はシナプス可塑性が担っている
扁桃体のエングラム細胞上の異なるシナプスが、記憶のアイデンティティを担っている可能性を検討するため、聴覚野のエングラム細胞と扁桃体のエングラム細胞の間のシナプスに長期抑圧(LTD)注12)を誘導し、その影響を調べました(図3)。2つの音恐怖条件付けを5時間間隔で行うにあたり、最初の7kHzの音に対して活動した聴覚野エングラム細胞をoChIEFで標識し、扁桃体に投射している神経終末の活動を光照射で操作してLTDを誘導しました(図3A)。その後、マウスは7kHzの音に対し低いすくみ反応を示し音恐怖記憶の減弱を示したのに対して(テスト3、図3B)、2kHzの音恐怖記憶は影響を受けず正常でした(テスト4、図3B)。この実験では、聴覚野の7kHzエングラム細胞と扁桃体の音恐怖エングラム細胞の間のシナプスのみにLTDを誘導していることから、この結果は記憶のアイデンティティはエングラム細胞上に存在する異なるシナプスの可塑性が担っていることを意味しています。
また、相補的な実験として、エングラム細胞間のシナプス特異的に長期増強(LTP)注12)を誘導しその影響を調べました(図4A)。2つの記憶を形成した後、それぞれの記憶を想起した直後に扁桃体にアニソマイシンとタットベクリンを投与して再固定化を阻害し、それぞれの音恐怖記憶に逆行性健忘を引き起こしました。その後、上記と同じ7kHzエングラム細胞と音恐怖エングラム細胞の間のシナプスに光遺伝学的にLTPを誘導すると、7kHzの音恐怖記憶は回復したのに対して、2kHzの音恐怖記憶は減弱したままでした(図4B)。
結論
以上の結果より、2つの記憶の相互作用はエングラム細胞の共有化で行われている一方、それぞれの記憶のアイデンティティはエングラム細胞上のシナプス可塑性を記憶ごとに使い分けることで担保されていることが初めて明らかになりました。
<今後の展開>
さまざまな記憶がアイデンティティを保ちながら、関連付けられていく仕組みを解明した今回の研究は、知識や概念を形成するヒトの精神活動の理解に向けての重要な一歩となります。記憶のアイデンティティの喪失は、PTSD(心的外傷後ストレス障害)を始めとする精神疾患や記憶障害を伴う認知症などに密接に関わっていることから、今回の研究成果はそれら多くの疾患の根本的な予防法や治療法の創出につながると期待されます。
<参考図>
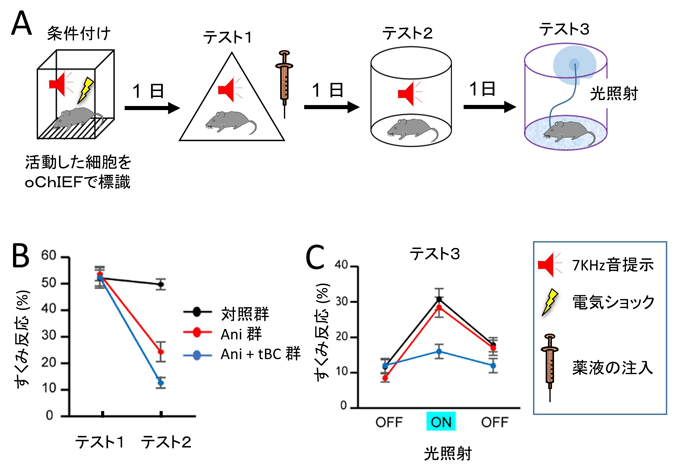
図1 逆行性健忘はエングラムの消失による
- A:逆行性健忘を引き起こす行動実験の手順。
- B:健忘後の音恐怖記憶のレベル。アニソマイシン投与群では、翌日のテスト2ですくみ反応が部分的に低下した。一方、タットベクリンを同時に投与した群は10パーセント程度のすくみ反応を示し、完全な逆行性健忘が誘導された。縦軸は、音を提示している時間のうちすくみ反応を示している時間の割合。マウスの無動状態をすくみ反応として自動判定している。恐怖条件付けされていないマウスのすくみ反応(無動時間)は約10パーセントである。
- C:健忘後に、光照射でエングラム細胞を人為的に活性化したときの音恐怖記憶のレベル。Ani+tBC群は、光照射の前後(OFF)でも光照射中(ON)のいずれでも恐怖反応を示さないが、対照群とAni群は光照射時(ON)に高いすくみ反応を示した。
Ani:アニソマイシン、tBC:タットベクリン
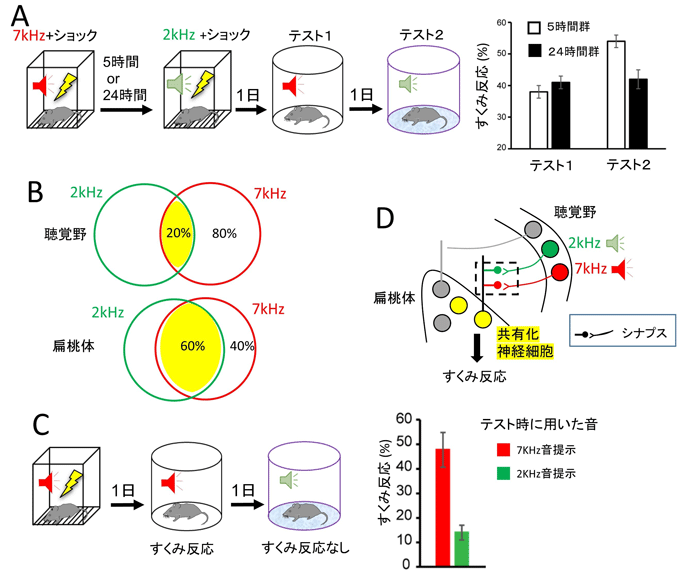
図2 エングラム細胞を共有しても記憶のアイデンティティは保たれている
- A:行動実験の手順とすくみ反応。5時間間隔で条件付けを行うと、2つの記憶が相互作用した。
- B:エングラム細胞群の共有化を示すベン図。2つの音恐怖が相互作用するとき、扁桃体においてはエングラム細胞群の共有化(60パーセント)が見られたが、聴覚皮質ではそれらは独立(20パーセント)していた。
- C:左側、7kHz音とショックによる条件付けの実験手順。右側、7kHz音でのみすくみ反応を示し、2kHz音ではすくみ反応を示さなかった。
- D:扁桃体と聴覚皮質における2つの音恐怖記憶に対するエングラム細胞の関係を模式的に表した。
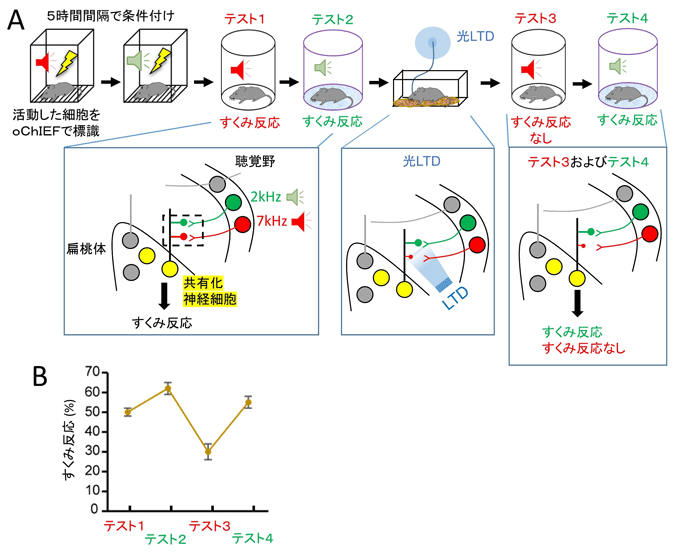
図3 シナプス特異的な可塑性が記憶のアイデンティティを担っている
光照射によるLTD実験
- A:実験手順(上段)と実験結果(下段)を示した模式図。
- B:各テスト時におけるすくみ反応。
条件付けにより、聴覚野の2kHzエングラム細胞および7kHzエングラム細胞と扁桃体の共有化エングラム細胞の間のシナプス伝達効率が上がるため、それぞれの音提示でマウスは高いすくみ反応を示した(テスト1、2)。光照射で7kHZ経路のシナプスにLTDを誘導すると、7kHZの音提示は扁桃体のエングラム細胞を活性化させることができないため、マウスはすくみ反応を示さなかった(テスト3)。一方で、2kHzの音提示は高いすくみ反応を示し、LTDによる影響は受けなかった(テスト4)。
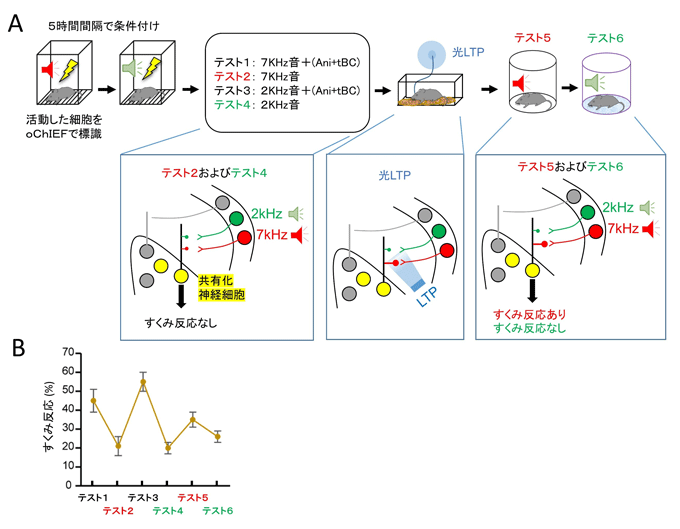
図4 シナプス特異的な可塑性が記憶のアイデンティティを担っている
光照射によるLTP実験
- A:実験手順(上段)と実験結果(下段)を示した模式図。
- B:各テスト時におけるすくみ反応。
条件付けにより、聴覚野の2kHzエングラム細胞および7kHzエングラム細胞と扁桃体の共有化エングラム細胞の間のシナプス伝達効率が上がるため、それぞれの音提示でマウスは高いすくみ反応を示した(テスト1、3)。7kHz(テスト1)および2kHz(テスト3)の音による恐怖記憶の想起後にAni+tBCを投与すると、どちらの記憶も逆行性健忘を示した(テスト2、4)。光照射で7kHz経路のシナプスにLTPを誘導すると、7kHzの恐怖記憶が蘇った(テスト5)。一方で、2kHzの恐怖記憶は健忘状態のままであった(テスト6)。
Ani:アニソマイシン、tBC:タットベクリン
<用語解説>
- 注1)記憶のアイデンティティ
- 記憶同士の間に関連付けが行われても、それぞれの記憶そのものは異なるものとしてアイデンティティを保っている。例えば、東京ドームで野球観戦した後に銀座のレストランでディナーを楽しんだ、というように連続して起きた出来事の記憶は関連付けて覚えているが、もちろん野球観戦とディナーは別々の出来事として記憶されている。
- 注2)エングラム細胞
- 記憶を担う神経細胞群のこと。記憶は脳内にある特定の神経細胞集団として符号化されて蓄えられる。学習時に活性化した特定の神経細胞のセットという形で脳の中に残った物理的な痕跡が「エングラム(痕跡)」である。何らかのきっかけでこの神経細胞集団が活動すると、その記憶が想起される。
- 注3)扁桃体
- 情動反応の処理を司る脳領域のこと。恐怖条件付けでは、音などの条件刺激(注6参照)の情報とショックなどの無条件刺激(注7参照)の情報が扁桃体で関連付けられて恐怖記憶を形成する。
- 注4)シナプス可塑性
- 経験に応じて神経細胞間のシナプスの伝達効率が変化(増強、あるいは減弱)し、その変化が長時間続くこと。細胞レベルでの学習・記憶の基礎過程であると考えられている。以下に記載されている長期抑圧(LTD)や長期増強(LTP)が代表例である。
- 注5)逆行性健忘
- 過去の記憶を思い出すことができない障害のことを指す。逆行性健忘は記憶そのものが失われたために生じるのか(エングラムの消失)、あるいは、エングラムは残っているが想起のプロセスに障害があるのかについては議論となっていた。
- 注6)再固定化の阻害
- 再固定化とは、脳内に保存された記憶が想起された後に、その記憶を再び固定し脳内に再保存するプロセスのことを指す。固定化(安定化)された記憶においても、その記憶は想起されると、一度不安定な状態となることがあり(不安定化)、脳内に安定した状態で再保存されるには、「再固定化」が必要である。再固定にはたんぱく質合成が必要であるため、想起時にたんぱく質合成を抑制すると再固定化が阻害され、逆行性健忘を引き起こす。
- 注7)条件刺激、無条件刺激
- 条件刺激:動物に対してそれ自体では恐怖反応などを誘導しない音、光、場所(文脈)などの刺激。
- 無条件刺激:電気ショックなどのように恐怖反応を引き起こす刺激。
これらを対提示することにより両者の関連性を学習し、条件刺激のみで恐怖反応を示すようになる。本研究の音恐怖条件付けでは、条件刺激としてブザー音、無条件刺激として足への電気ショックを行った。
- 注8)アニソマイシン
- たんぱく質合成阻害剤。記憶の想起時にアニソマイシンを投与すると、記憶の再固定化が阻害され逆行性健忘を引き起こす。多くの場合、想起に伴う記憶の不安定化は部分的であるため、部分的な逆行性健忘を引き起こす。
- 注9)タットベクリン
- たんぱく質分解系のオートファジー活性を促進するペプチド。記憶の想起に伴い、エングラム細胞のオートファジーが活性化され、シナプス膜に存在するグルタミン酸受容体を分解する。その結果、シナプス伝達が減衰し記憶の減弱化を引き起こす。タットベクリンは減弱化効果を促進する。
- 注10)oChIEF(オチーフ)
- 光照射により活性化される非選択的陽イオンチャネルである。その際、oChIEF発現細胞は脱分極応答を示し活性化されるため、光照射依存的かつ標的細胞特異的に神経活動を誘導することができる。ミリ秒オーダーの非常に早い光反応時間を持つため、100Hzなどの高頻度の光刺激に対しても適している。
- 注11)catFISH法(キャットフィッシュ法)
- 蛍光を利用しmRNAの局在を調べるFISH法を利用して、神経細胞が活動したタイミングを同定する技術。Arc mRNAは神経活動5分後に、Homer1a mRNAは神経活動30分後に核内に発現が確認される。この時間差を利用し、30分間隔で2回のイベントを行った直後の脳サンプルを解析することで、神経細胞がいつ活動したのかを調べることができる。
- 注12)長期抑圧(LTD)、長期増強(LTP)
- シナプスの伝達効率は、シナプス前後の神経細胞の活動パターンにより、長期間にわたり増強したり(LTP)、減弱したりする(LTD)。本研究では、光遺伝学的手法により、シナプス前細胞の神経終末に発現したoChIEFに光照射することで、人為的にLTDやLTPを誘導している。1Hzの連続した光照射でLTDを、100Hzの光照射でLTPを誘導することができる。
<論文情報>
| タイトル |
“Synapse-specific Representation of the Identity of Overlapping Memory Engrams”
(シナプス特異性による記憶のアイデンティティの保持) |
| doi |
10.1126/science.aat3810 |
<お問い合わせ先>
<研究に関すること>
井ノ口 馨(イノクチ カオル)
富山大学 大学院医学薬学研究部(医学) 教授
〒930-0194 富山県富山市杉谷2630
Tel:076-434-7225 Fax:076-434-5014
E-mail:
<JST事業に関すること>
川口 哲(カワグチ テツ)
科学技術振興機構 戦略研究推進部 ライフイノベーショングループ
〒102-0076 東京都千代田区五番町7 K’s五番町
Tel:03-3512-3524 Fax:03-3222-2064
E-mail:
<報道担当>
科学技術振興機構 広報課
〒102-8666 東京都千代田区四番町5番地3
Tel:03-5214-8404 Fax:03-5214-8432
E-mail:
富山大学 総務部 総務・広報課
〒930-8555 富山県富山市五福3190
Tel:076-445-6028 Fax:076-445-6063
E-mail: