ポイント
- 根粒菌と共生して大気中の窒素を利用するマメ科植物が、土壌中の窒素栄養が豊富な場合に根粒共生を抑制する転写因子NRSYM1を発見しました。
- 長年未解明だった根粒共生に伴うエネルギー消費を抑える仕組みに関する初めての発見です。
- 窒素肥料の効率的な管理など、マメ科作物の持続可能な農業実現への貢献が期待されます。
筑波大学 生命環境系 寿崎 拓哉 准教授(つくば機能植物イノベーション研究センター)、西田 帆那(総合研究大学院大学 大学院生)らの研究グループは、基礎生物学研究所、東京理科大学との共同研究により、NITRATE UNRESPONSIVE SYMBIOSIS 1(NRSYM1)と名付けた転写因子注1)が土壌中の窒素栄養に応答して根粒共生注2)を抑制することを発見しました。
マメ科植物は土壌中の根粒菌と根粒共生を行い、根粒菌の窒素固定注3)によって、窒素栄養が乏しい環境でも大気中の窒素を利用しています。ただし、宿主植物は光合成産物を根粒菌にエネルギー源として供給しなければいけません。そこで宿主植物は、窒素栄養が土壌中に十分に存在する場合には、根粒共生を抑制し、共生に伴うエネルギーの消費を抑えることが知られていましたが、その制御に関わる分子機構の理解は進んでいませんでした。
本研究グループは、マメ科のモデル植物であるミヤコグサ注4)を用いた突然変異体のスクリーニングによって変異体nrsym1を単離しました。このnrsym1変異体は、野生型植物では根粒共生を抑制するような高濃度の硝酸を添加した条件でも、共生を抑制することができませんでした。このことから変異の原因遺伝子であるNRSYM1遺伝子は、根粒の数を制御するペプチド性シグナル因子CLE-RS2の生産を誘導していて、高濃度の硝酸に応答した根粒共生の抑制を制御する因子であることがわかりました。さらにNIN-LIKE PROTEIN (NLP)と呼ばれるタイプの転写因子をコードすることを特定しました。
硝酸による根粒共生の抑制において中心的な役割を果たすと考えられるNRSYM1転写因子の発見は、植物がどのように土壌中の窒素栄養環境に応答して根粒共生を適切に制御しているかという長年解明できなかった問題の理解に向けた重要な基礎的知見を提供するものです。また、NRSYM1の解析から得られた知見は、大豆栽培における肥料管理など、根粒共生を利用した持続可能な農業の実現のためにも役立つことが期待されます。
本研究の成果は、2018年2月5日(日本時間)付で「Nature Communications」で公開される予定です。
本研究は、科学研究費補助金 新学術領域研究「植物の成長可塑性を支える環境認識と記憶の自律分散型統御システム」の助成によって実施され、科学技術振興機構(JST) 戦略的創造研究推進事業 総括実施型研究(ERATO)「野村集団微生物制御プロジェクト」の一環で行われました。
<研究の背景>
窒素は、植物が生きる上で必須の栄養素です。しかし、窒素は大気中にたくさん存在するにもかかわらず、多くの植物はその窒素を直接利用することができません。そのため、土壌中に存在する窒素栄養(主に硝酸やアンモニア)を利用して生きています。ところがマメ科植物は、土壌中の窒素栄養が乏しい環境でも生育することが可能です。それは、根に根粒と呼ばれるこぶ状の器官を形成し、その中に、大気中の窒素を植物が利用できる形に変換してくれる根粒菌(窒素固定細菌)を共生させているからです。しかし、根粒菌と共生するにあたっては、植物が自ら生産した光合成生産物を、窒素固定反応などのエネルギー源として根粒菌へ供給しなければなりません。そのため、宿主植物は光合成産物を消費しすぎることで成長に悪影響を及ぼさないように、自身の根に作る根粒の数を適切に保つ必要があり、そのための仕組みを備えています。また、土壌中に窒素栄養が豊富に存在する場合には、そちらを優先的に利用することができるので、エネルギーのかかる根粒菌との共生をやめて光合成産物の消費を抑えています。この土壌中の窒素栄養に応答した根粒共生の抑制は、古くから知られた現象であるものの、その制御機構についての理解はあまり進んでいませんでした。
<研究内容と成果>
本研究では、マメ科のモデル植物であるミヤコグサを用いて、窒素栄養(硝酸)が豊富に存在する土壌でも根粒を形成するような突然変異体のスクリーニングを行い、新しい変異体を単離してnitrate unresponsive symbiosis 1 (nrsym1)と名付けました。nrsym1変異体は、窒素栄養を含まない土壌では野生型と同様の根粒共生を示します。高濃度の硝酸を添加した場合、野生型の植物は根粒を作らなくなりますが、nrsym1変異体は窒素栄養を含まない条件とほとんど変わらない成熟した根粒を形成しました(図1)。したがって、NRSYM1遺伝子は、高濃度の硝酸に応答して引き起こされる根粒共生を抑制する因子であることがわかりました。nrsym1変異体の原因遺伝子を特定したところ、NIN-LIKE PROTEIN (NLP)と呼ばれるタイプの転写因子をコードしていることが明らかになりました。シロイヌナズナなどの研究から、NLPは植物が硝酸に応答してさまざまな遺伝子の発現を制御する硝酸応答の中心的な役割を担う因子であることが示唆されています。
これまでの研究から、宿主植物が備えている根粒の数を適切に保つ仕組みは、根と地上部を介した全身的なシグナル伝達(Autoregulation of nodulation:AON注5)と呼ばれる)によって制御されていることが明らかにされています。しかもこのAONは、窒素栄養に応答した根粒共生の抑制にも関わっている可能性が示唆されていました。野生型植物では、AONにおいて根粒菌の感染を根から地上部へ伝えるためのシグナルとして働くCLEペプチド遺伝子の1つであるCLE-RS2遺伝子の発現が、根粒菌の感染だけでなく硝酸によっても強く誘導されます。一方、nrsym1変異体では、硝酸を添加してもCLE-RS2遺伝子の発現は誘導されないため、硝酸に応答したCLE-RS2遺伝子の発現をNRSYM1転写因子が制御している可能性が考えられました。そこで、CLE-RS2遺伝子がNRSYM1の直接の標的遺伝子であるかを検証した結果、NRSYM1転写因子は硝酸に応答してCLE-RS2遺伝子の発現を直接誘導し、AONと同様の機構を用いて全身的に根粒の数を制御していることが明らかになりました(図2)。
また、NRSYM1タンパク質の細胞内局在を調べてみると、NRSYM1タンパク質は硝酸添加時には核に局在し、無窒素の条件では核への局在がほとんど見られなくなることがわかりました(図3)。したがって無窒素条件では核の外にいるNRSYM1が、硝酸に応答して核に移行し、標的遺伝子の発現を誘導することで、硝酸(窒素栄養)が豊富に存在する場合には根粒共生を抑制する可能性が示唆されました。
<今後の展開>
本研究によって、NRSYM1転写因子が硝酸に応答した根粒共生制御の中心的な働きを担うことが明らかになりました。NRSYM1は、さまざまな下流因子を使い分けることで根粒共生を多面的に制御していると予想されます。今後はCLE-RS2遺伝子以外のNRSYM1標的遺伝子を同定することで、硝酸に応答した根粒共生の制御機構の全容解明に迫っていきたいと考えています。窒素栄養に応答した根粒共生の抑制の仕組みを理解することは、窒素肥料と根粒共生による窒素源の獲得の両立のために重要であり、NRSYM1の研究から得られた知見は、マメ科作物の効率的な肥料管理など、持続可能な農業の実現に貢献する可能性が期待できます。
<参考図>
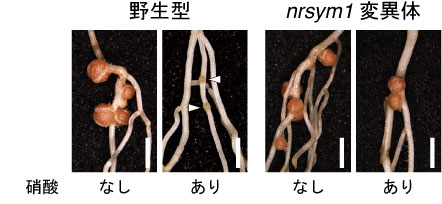
図1
野生型植物と、窒素栄養が豊富な状況でも根粒を形成するnrsym1変異体の根粒共生。矢じりは未熟な根粒を示す。スケールバー:2mm
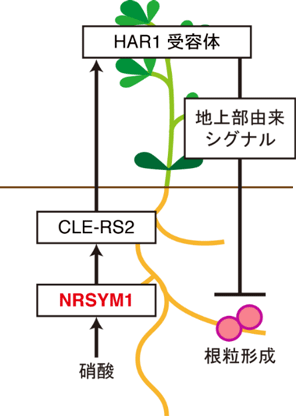
図2 NRSYM1転写因子によるAONを介した全身的な根粒数の制御機構のモデル図
土壌中の硝酸に応答したNRSYM1は根においてCLE-RS2遺伝子の発現を直接誘導する。地上部へと移動したCLE-RS2が受容体(HAR1)によって受容された後、地上部由来シグナルが生産されて根へと移動し、根粒形成を抑制する。
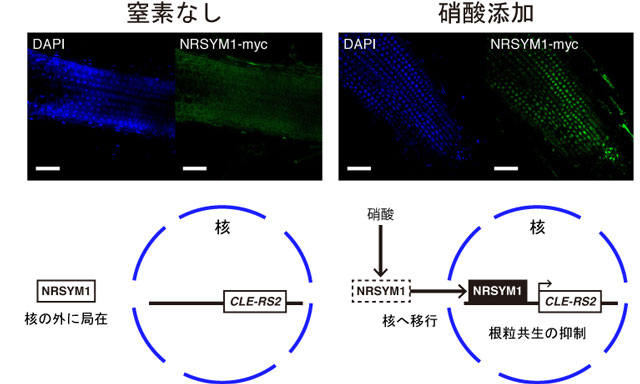
図3 NRSYM1タンパク質の細胞内局在と遺伝子発現制御モデル
核の位置をDAPI染色(青色)、NRSYM1タンパク質の局在を抗体免疫染色(緑色)によってそれぞれ可視化している。スケールバー:50μm
無窒素の条件では核の外に局在しているNRSYM1は、硝酸添加時には硝酸に応答して核へと移行し、標的遺伝子の発現を誘導することにより根粒共生を抑制する。
<用語解説>
- 注1)転写因子
- DNA配列に特異的に結合し、標的となる遺伝子の発現を調節する機能を持つタンパク質の総称。
- 注2)根粒共生
- マメ科植物は土壌中に存在する根粒菌との相互作用によって根に根粒と呼ばれる器官を形成する。根粒の中には根粒菌が共生しており、大気中の窒素を固定する。宿主植物は根粒菌から固定された窒素源を受けとる代わりに、エネルギー源として光合成産物を根粒菌に与える。
- 注3)窒素固定
- 大気中の窒素を植物が利用できるアンモニアの形に変換する反応。根粒共生においては、根粒に細胞内共生する根粒菌が行っている。
- 注4)ミヤコグサ
- マメ科のモデル植物。ゲノムサイズが比較的小さく、形質転換法が確立されているなどの理由から遺伝学実験に適しており、植物微生物相互作用の研究材料として広く利用されている。
- 注5)AON (Autoregulation of nodulation)
- マメ科植物が備える根粒の数を適切に保つための全身的な根粒形成の抑制機構。根粒菌が感染すると、根でシグナル分子(CLEペプチド)が生産されて地上部へ移動し根粒菌が感染したことを伝える。地上部で働くシグナルの受容体(HAR1)によりその情報が受けとられると、今度は地上部から根へと根粒形成を抑制するためのシグナルが送られ、根では新しい根粒形成が抑えられる。宿主植物はこの仕組みを用いて、根粒を過剰に形成してしまうことを防いでいる。
<論文情報>
| タイトル |
“A NIN-LIKE PROTEIN mediates nitrate-induced control of root nodule symbiosis in Lotus japonicus”
(ミヤコグサ NIN-LIKE PROTEIN は硝酸に応答して根粒共生を制御する) |
| 著者名 |
Hanna Nishida, Sachiko Tanaka, Yoshihiro Handa, Yuki Sakamoto, Sachihiro Matsunaga, Shigeyuki Betsuyaku, Kenji Miura, Takashi Soyano, Masayoshi Kawaguchi, Takuya Suzaki. |
| doi |
10.1038/s41467-018-02831-x |
<お問い合わせ先>
<研究に関すること>
寿崎 拓哉(スザキ タクヤ)
筑波大学 生命環境系、つくば機能植物イノベーション研究センター 准教授
〒305-8572 茨城県つくば市天王台1-1-1
Tel:029-853-4674
E-mail:
<JST事業に関すること>
大山 健志(オオヤマ タケシ)
科学技術振興機構 研究プロジェクト推進部
〒102-0076 東京都千代田区五番町7 K’s五番町
Tel:03-3512-3528
E-mail:
<報道担当>
科学技術振興機構 広報課
〒102-8666 東京都千代田区四番町5番地3
Tel:03-5214-8404 Fax:03-5214-8432
E-mail: