名古屋大学 大学院生命農学研究科(研究科長:川北 一人)の森田(寺尾) 美代(モリタ(テラオ) ミヨ) 教授と古谷 将彦(フルタニ マサヒコ) 特任助教、西村 岳志(ニシムラ タケシ) 特任助教は、奈良先端科学技術大学院大学の田坂 昌生(タサカ マサオ) 名誉教授、大阪大学の田中 博和(タナカ ヒロカズ) 助教、埼玉大学の豊田 正嗣(トヨタ マサツグ) 准教授らとの共同研究で、シロイヌナズナの根、胚軸、茎すべての器官の重力屈性を制御する遺伝子ファミリー注1)を見いだしました。また、その機能は重力感受細胞注2)における重力刺激に応じたオーキシン輸送制御注3)であることを明らかにしました。
植物は多くの枝や側根を持っており、これらは重力方向に対して一定の角度を保って成長することで、枝振りや根の張りにおいて特徴ある植物構造(プラントアーキテクチャ)をとります。プラントアーキテクチャは、植物が地上および地下部において資源取得を最適化するための機構の1つと考えられ、育種上重要な形質です。今回の研究では、主茎や主根の重力屈性と同じ重力情報伝達機構が、プラントアーキテクチャを決める側枝や側根の角度制御にも使われていることが分かりました。さらに研究を進めることで、将来的には有用植物の育種やプラントアーキテクチャを制御する成長調節剤の開発につながると期待されます。
この研究成果は、平成29年8月1日付(米国東部時間午前10時)米国科学雑誌 「The Plant Cell」オンライン版に掲載されます。 この研究は、科学技術振興機構 戦略的創造研究推進事業 CREST、日本学術振興会NEXT programなどの支援のもと行われました。
<研究背景と内容>
植物の根は地中に向かって、茎は空に向かって成長します。これは、植物が重力の方向を感じ取り、根を水分や栄養分が豊富な地中へ、地上部を光合成や生殖に有利な上方へと体の各器官を配置する重要な環境応答の1つで、重力屈性といいます。重力屈性を示す根や茎などの器官には、重力方向の変化(器官の傾き)を受容する重力感受細胞があります。この細胞の色素体はデンプンを高度に蓄積してアミロプラスト注4)となり、細胞質よりも比重が高く重力方向に沈降しますが、これが重力方向の認識に重要であることが知られています。重力方向が認識された後、その器官内で成長を調節する植物ホルモンであるオーキシン濃度に差が生じ、器官が屈曲することが示されています(図1)。つまり、重力感受細胞で受容された重力刺激に従って器官内でのオーキシン分布が制御されますが、感受細胞内での情報伝達の仕組みは明らかにされていませんでした。今回、モデル植物であるシロイヌナズナを用いて、植物特有の遺伝子LZY(LAZY1-like)ファミリーが、重力感受細胞内で重力刺激に応じたオーキシン分布制御を行うことを明らかにしました。
この研究では、まず重力感受細胞分化不全により花茎の重力屈性能を失った突然変異体eal1と野生型の花茎からRNAを抽出し、マイクロアレイ注5)という手法を用いてeal1で発現が低下する遺伝子を探しました。これらの遺伝子の中には、感受細胞で発現して重力情報伝達に機能する遺伝子が含まれることを期待しました。多くの候補遺伝子の中に、イネ地上部の重力屈性異常変異体の原因遺伝子であるLAZY1遺伝子に低い相同性を示す遺伝子が2つ見つかりました。これらをLZY1、LZY2と名付け、LZY2と相同性の高いLZY3と共に、シロイヌナズナにおいて機能解析を行いました。単独および多重変異体を作成し、重力屈性能を解析した結果、花茎、胚軸、根それぞれの器官で貢献度は異なるものの、LZY1、LZY2、LZY3は重力屈性では冗長的に働くことが分かりました。次にこれらの遺伝子が期待通り重力感受細胞で発現するかを調べたところ、LZY1、LZY2、LZY3は花茎、胚軸において重力感受細胞である内皮細胞で発現し、LZY2、LZY3は根の重力感受細胞であるコルメラ細胞で発現していることが分かりました(図2)。また、重力感受細胞特異的な遺伝子発現をさせるプロモーターの制御下においたLZY遺伝子が、変異体の重力屈性異常を回復させたことから、LZY遺伝子は重力感受細胞で機能することを明らかにしました。重力屈性能を失った多重変異体では、重力感受細胞は正常に形成され、アミロプラストの重力方向への沈降も正常に起こりますが、重力刺激に応じた器官内でのオーキシン偏差分布が生じないことを見いだしました。従って、LZY遺伝子の機能は、アミロプラスト沈降後、その情報をオーキシン偏差分布へとつなげる感受細胞内での情報伝達であると言えます。
植物は多くの枝や側根を持っており、これらは重力方向に対して一定の角度を保って成長する傾斜重力屈性もしくは横重力屈性と呼ばれる性質を示します。シロイヌナズナの側枝や側根も斜めの角度を保ちますが、lzy多重変異体では側枝や側根が重力とは逆の方向に斜めの角度を保つという性質を示すことが分かりました(図3)。重力感受細胞特異的にLZY遺伝子を発現させると、正常な角度に戻ることから、主茎や主根の重力屈性と同じ重力情報伝達機構が側枝や側根の角度制御にも使われていることが分かりました。側根の角度制御におけるLZY遺伝子の役割をさらに詳しく調べ、重力感受細胞で発現するオーキシン排出輸送体PIN3の根冠での発現パターンを重力方向に偏らせる機能を持ち、これによってオーキシンを重力方向に偏差分布させる(図4)ことで、水平方向に伸長している若い側根の伸長角度を重力側に傾けるという新しいモデルを提示しました。
<成果の意義>
重力屈性と側生器官の伸長角度制御は、植物器官の空間配置によるプラントアーキテクチャ(植物構造:いわゆる枝振り、根の張りや樹冠)をつかさどる要因の1つで、草丈や枝数と同様に育種上重要な形質です。イネやトウモロコシにおいては葉の角度を上に向ける形質が密植や光合成効率の改善につながり、生産量を増大させました。このような農業的重要度に比して、その分子メカニズムの学術的な研究は少なく、その理解は進んでいませんでした。今回の研究で得られた成果は、これまで未知であった重力屈性における重力情報伝達の分子機構の理解を大きく前進させました。また、この遺伝子ファミリーの研究をさらに進めることで、将来的には有用植物の育種やプラントアーキテクチャを制御する成長調節剤の開発につなげることができると期待されます。
<参考図>
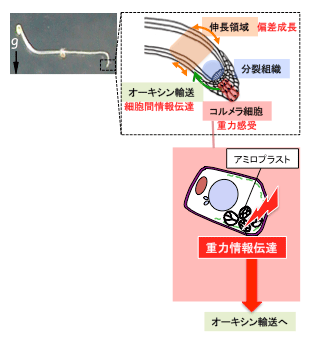
図1 シロイヌナズナの主根における重力屈性反応
重力感受細胞内のアミロプラスト沈降による重力感受から、器官の偏差成長に至る情報の流れを図示した。今回は、感受細胞内での重力情報伝達に関わる遺伝子を見いだした。
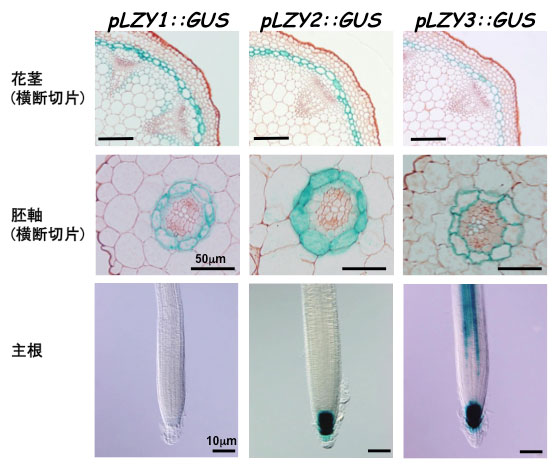
図2 LZY1、LZY2、LZY3は主に重力感受細胞で発現している
各遺伝子のプロモーター領域(pLZY)にGUS(β-グルクロニダーゼ)遺伝子を連結したものを野生型植物に導入した形質転換体を用いた。各遺伝子が発現する組織が、GUSの働きにより青色に染色される。花茎および胚軸では内皮が、主根では根端のコルメラがよく染色されている。
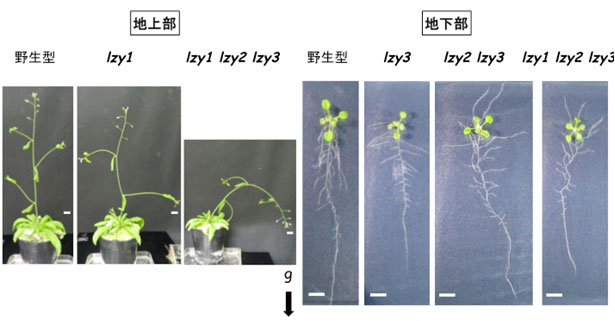
図3 LZYの機能欠損は主軸の重力屈性に加え、側枝および側根の伸長方向に影響する
lzy1、lzy2、lzy3三重変異体の側枝は下方に、側根は上方に伸長した。地上部は播種後約6週の植物体、地下部は播種後約2週の植物体。gと矢印は重力の方向を示す。
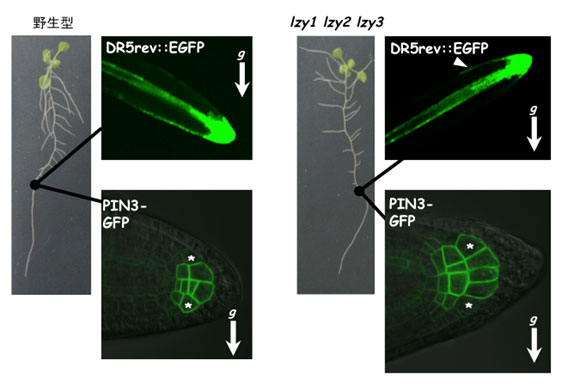
図4 側根端におけるオーキシンシグナルと側根コルメラにおけるオーキシン輸送体の発現
オーキシン量の増加に応じて緑(GFP)のシグナルが濃くなるオーキシンシグナルマーカーDR5rev::EGFPと、コルメラ細胞で発現しオーキシン輸送に関わるPIN3にGFPを融合した遺伝子PIN3-GFPを導入した形質転換体を用いて、若い側根を観察した。野生型では両シグナルは重力方に偏るが、lzy1、lzy2、lzy3三重変異体では逆方向への偏りが見られる。アスタリスクはコルメラの対称な位置を示している。gと矢印は重力の方向を示す。
<用語解説>
- 注1) 遺伝子ファミリー
- 同一の祖先遺伝子に由来し、配列が互いに似ている一群の遺伝子をいう。
- 注2) 重力感受細胞
- 重力屈性において重力方向の変化を感受する細胞。細胞内に重力方向に移動することができるアミロプラストを含む。
- 注3) オーキシン輸送制御
- オーキシンは細胞伸長、細胞極性や器官の軸性の形成と維持などさまざまな生理・発生現象を制御する植物ホルモンである。オーキシン輸送体PINファミリータンパク質によってオーキシンは組織内を特定の方向に輸送される。オーキシン輸送の制御は、植物器官内のオーキシンの分布パターンの決定要因のひとつであり、発生や環境応答に大きな影響を与える。
- 注4) アミロプラスト
- 植物細胞に含まれる色素体は、細胞の性質に合わせて葉緑体、有色体、白色体などさまざまな形態に分化する。一般にアミロプラストは、デンプンを高度に蓄積した白色体のことを指す。重力感受細胞に含まれるアミロプラストは、細胞質に比べて高比重で、重力方向に沈降する性質を持ち、重力感受において平衡石の役割を持つと考えられている。
- 注5) マイクロアレイ
- 数万にのぼる遺伝子を一度に検出する方法で、網羅的な遺伝子発現解析によく用いられる。本研究では、この方法を用いて野生型とeal1変異体との間で網羅的に遺伝子発現量の比較を行った。
<論文情報>
| タイトル |
“Arabidopsis LAZY1 family plays a key role in gravity signaling within statocytes and in branch angle control of roots and shoots.” |
| 著者名 |
Masatoshi Taniguchi, Masahiko Furutani, Takeshi Nishimura, Moritaka Nakamura, Toyohito Fushita, Kohta Iijima, Kenichiro Baba, Hirokazu Tanaka, Masatsugu Toyota, Masao Tasaka, and Miyo Terao Morita |
| 掲載誌 |
The Plant Cell |
| doi |
10.1105/tpc.16.00575 |
<お問い合わせ先>
<研究に関すること>
森田(寺尾) 美代(モリタ(テラオ) ミヨ)
名古屋大学 大学院生命農学研究科 教授
Tel:052-789-4123 Fax:052-789-5746
E-mail:
<JST事業に関すること>
川口 哲(カワグチ テツ)
科学技術振興機構 戦略研究推進部 ライフイノベーショングループ
Tel:03-3512-3524 Fax:03-3222-2064
E-mail:
<広報担当>
名古屋大学 総務部 広報渉外課
Tel:052-789-2699 Fax:052-788-6272
E-mail:
科学技術振興機構 広報課
〒102-8666 東京都千代田区四番町5番地3
Tel:03-5214-8404 Fax:03-5214-8432
E-mail: