ポイント
- マウス多能性幹細胞から精子幹細胞様細胞の試験管内での誘導に成功。
- 精子幹細胞様細胞は成体の精巣内で精子に分化し、健常な子孫を産生。
- 精子幹細胞におけるDNAのメチル化異常が精子形成不全につながることを発見。
京都大学 大学院医学研究科の斎藤 通紀 教授[兼 科学技術振興機構(JST) ERATO斎藤全能性エピゲノムプロジェクト 研究総括、京都大学物質-細胞統合システム拠点(iCeMS=アイセムス) 主任研究者、京都大学 iPS細胞研究所 研究員]、同研究科の石藏 友紀子 特定研究員らは、マウス多能性幹細胞注1)(ES細胞)から、試験管内にて精子幹細胞様細胞注2) 注3)およびその長期培養株Germline stem cell-like cells(GSCLCs)を誘導することに成功しました(図1)。このGSCLCsは、生殖細胞を欠損する成体マウスの精巣中で精子に分化し、健常な子孫を生み出すことができました。
本研究グループは、これまで、マウス多能性幹細胞から始原生殖細胞様細胞注4) 注5)を誘導し、それらを、生殖細胞欠損マウス新生仔(生後7日齢)の精巣に移植することで精子を得、さらには健常な産仔を得ることに成功してきました。オスの生殖細胞発生過程を試験管内で再構成する研究において、次の重要な目標は精子幹細胞を誘導することです。精子幹細胞は、成体で常に精子を産出する生殖細胞系列で唯一の幹細胞です。
本研究では、マウス多能性幹細胞(ES細胞)から誘導した始原生殖細胞様細胞を、胎仔(胎齢12.5日齢)の生殖巣注6)体細胞と共に凝集させて作製した「再構成精巣」(図2)を培養することにより、始原生殖細胞様細胞から精子幹細胞に似た細胞を分化させ、これをGSCLCsとして試験管内で4ヶ月以上長期培養することに成功しました。さらに、このGSCLCsを、生殖細胞欠損マウスの新生仔(生後7日齢)および成体(生後8週齢)の精巣に移植したところ、その一部が両精巣中で精子に分化し、健常な子孫を生み出すことができました(図3)。この結果は、始原生殖細胞様細胞が長期間培養できないこと、新生仔精巣内でしか精子に分化できないという、これまでの2つの課題を解決しました。さらに本研究では、精子幹細胞形成過程におけるDNAのメチル化注7)制御異常が精子形成不全につながることを発見しました。今後、精子幹細胞の形成メカニズムの解明、DNAのメチル化異常に起因する疾患の発症メカニズムの解明、ヒト始原生殖細胞様細胞からヒト精子幹細胞様細胞を誘導する方法論の開発などに貢献すると期待されます。
本研究成果は、JST 戦略的創造研究推進事業の一環として行われ、2016年12月6日(米国東部時間)に米国科学誌『Cell Reports』のオンライン速報版で公開されます。
本成果は、以下の事業・研究プロジェクトによって得られました。
戦略的創造研究推進事業 総括実施型研究(ERATO)
| 研究プロジェクト |
「斎藤全能性エピゲノムプロジェクト」 |
| 研究総括 |
斎藤 通紀(京都大学 大学院医学研究科 教授) |
| 研究期間 |
平成23年度~平成28年度 |
<研究の背景>
生殖細胞は、哺乳類の体を構成する細胞の中で、次世代へと受け継がれ、新たな個体を作り出すことが可能な唯一の細胞です。生殖細胞系列の分化過程や、生殖細胞に特徴的なDNAのメチル化を含むエピゲノム情報の再構成注8)メカニズムを解明することは、不妊の原因究明や世代を経たエピゲノム情報の伝達メカニズムの理解につながります。生殖細胞分化の重要な過程の多くは、胎児成長の過程で行われます。しかし、胎児の生殖細胞は、その細胞数の少なさやサイズが小さいことによる扱いづらさから、解析に困難を伴います。そのため、多能性幹細胞から生殖細胞系列の細胞を試験管内で誘導する試みが、四半世紀にわたって行われてきました。近年、多能性幹細胞から精子や卵子の元となる始原生殖細胞を誘導する手法が確立され、それに続きオス、メス各々について配偶子分化過程の再現を目指す研究がなされてきました。メスについては、多能性幹細胞から始原生殖細胞を経て卵子を試験管内で誘導する手法が報告されています。一方オスについては、多能性幹細胞から始原生殖細胞を経て、精子の前段階の細胞である、精子幹細胞を誘導することが目標の一つとされてきました。精子幹細胞は、生涯にわたり精子を産出する細胞で、成体の精巣内にわずかしか存在せず、生殖細胞系列で唯一の幹細胞といわれています。これまで、精子幹細胞の長期培養株(GS細胞注9))の樹立方法は研究されていました。しかし、精子幹細胞が、始原生殖細胞から分化誘導されるメカニズムには不明な点が多く、多能性幹細胞から始原生殖細胞様細胞を経て、精子幹細胞を試験管内で誘導するシステムの確立が待たれていました。
<研究手法・成果>
本研究グループは、これまで多能性幹細胞から始原生殖細胞様細胞を試験管内で誘導する手法を確立してきました。オスにおいて、始原生殖細胞は胎齢12.5日齢までに将来精巣の元となる生殖巣体細胞に囲まれ、前精原細胞注10)と呼ばれるようになります。その後、前精原細胞は、出生5日齢頃に、精原細胞注11)および精子幹細胞へと分化します。本研究では、始原生殖細胞が前精原細胞となる時点の細胞環境に注目し、マウスES細胞から誘導した始原生殖細胞様細胞を、マウス胎仔(胎齢12.5日齢)の生殖巣体細胞と共に凝集させて「再構成精巣」を作製し、精子幹細胞への分化が誘導される培養条件を検討しました。その結果、始原生殖細胞様細胞が、精子幹細胞と同等の特性を示す細胞に分化する、培養方法と培養期間を決定しました。次に、この細胞を培養したところ、生体由来のGS細胞と同様に増殖し、4ヶ月以上の長期間の培養が可能であることが確認されました。我々は、この細胞株をGermline stem cell-like cells(GSCLCs)と命名しました。さらにGSCLCsを、生殖細胞欠損マウスの新生仔(生後7日齢)および成体(生後8週齢)の精巣に移植した結果、一部が精子まで分化し、得られた精子と卵子を顕微授精させると健常な産仔が得られることも確認しました。
一方で、樹立したGSCLCsがマウスの精巣中で精子まで分化する効率は、生体由来のGS細胞より低効率(20%程度)であることが分かりました。その要因を探るため、GSCLCsとGS細胞において、転写産物注12)とDNAのメチル化状態を比較しました。転写産物を解析したところ、GSCLCsではGS細胞と非常によく似た遺伝子発現パターンを示していましたが、精子幹細胞の分化に重要な一部の遺伝子の発現が、GSCLCsでは抑制される傾向にあることが分かりました。また、GSCLCsのゲノムには生体由来の精子幹細胞に比べて過剰に高いメチル化を示す領域が存在しており、それらの領域には精子に分化するために必要な遺伝子が含まれていました。以上から、転写産物の低発現と過剰メチル化に相関があることが示唆されました。つまり、試験管内で精子幹細胞へと分化させる過程で付与された過剰なメチル化が、精子分化に必要な遺伝子の発現を妨げるために、精子分化の効率が低くなることが示唆されました。これらの結果は、始原生殖細胞が精子幹細胞に分化する過程で起こるエピゲノム情報の再構成が、その後の精子分化に重要であることを示しています。
<波及効果、今後の予定>
本研究は、マウス多能性幹細胞から始原生殖細胞様細胞を経てGSCLCsを試験管内で誘導し、精子および健常な産仔を生み出すことに成功した初めての研究成果です。また、精子幹細胞形成過程におけるエピゲノム制御に異常が起こると、精子形成不全が起こる可能性が示唆されました。本研究で確立した培養システムと得られた知見は、男性不妊や、代謝疾患や精神疾患を含むエピゲノム異常症、遺伝病発症の原因究明に役立つことが期待されます。また、本研究は、ヒト始原生殖細胞様細胞からヒト精子幹細胞様細胞を誘導する方法論の開発に貢献すると期待されます。今後は、より質の高い培養システムの確立や、世代を超えたエピゲノム情報継承メカニズムの解明に向けて研究を進める予定です。
<参考図>
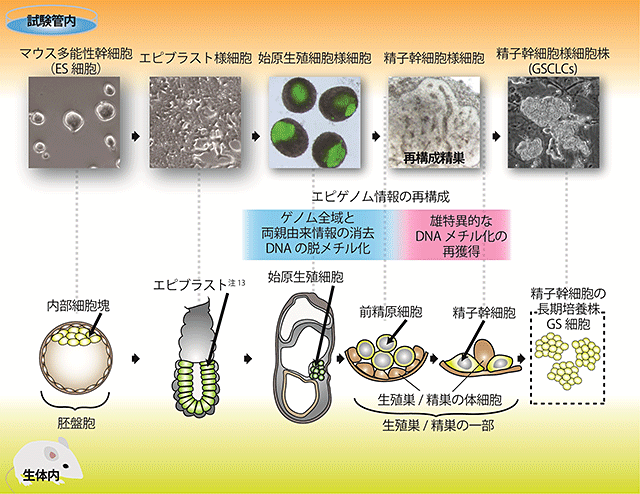
図1 マウス多能性幹細胞から精子幹細胞様細胞株(GSCLCs)を試験管内で誘導する概略図
- 上部:試験管内にて、マウス多能性幹細胞(ES細胞)から誘導した始原生殖細胞様細胞(緑色)と、オスの胎仔の生殖巣体細胞から作った再構成精巣の中で、始原生殖細胞様細胞は精子幹細胞様細胞へと分化した。そこから長期培養株であるGSCLCsを樹立した。その過程で、エピゲノム情報の再構成を試験管内にて一部再現した。
- 下部:生体内でのオスの生殖細胞分化過程を、試験管内での分化と対応させた。受精卵は発生が進むと胚盤胞となり、内部細胞塊ができる。そこから、エピブラスト注13)を経て、始原生殖細胞が出現する。その後、始原生殖細胞はオスの生殖巣体細胞に囲まれ、前精原細胞を経て、精原細胞や精子幹細胞へと分化する。精子幹細胞を長期培養すると、GS細胞となる。
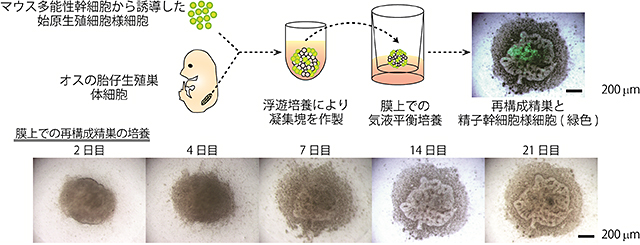
図2 再構成精巣の作製方法
再構成精巣は、始原生殖細胞様細胞(緑色)と、オスの胎仔(胎齢12.5日齢)生殖巣の体細胞を共に浮遊培養させて凝集塊を作り、それらを膜上で培養することで作製する。精巣に特徴的な管構造が自発的に再構成され、始原生殖細胞様細胞(緑色)が管構造内で分化する。
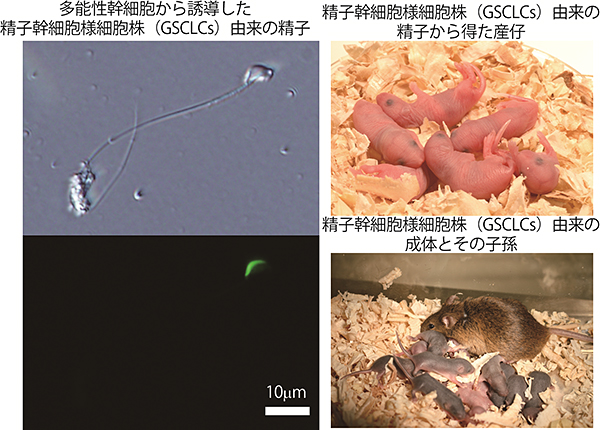
図3 マウス多能性幹細胞(ES細胞)から誘導したGSCLCsより得られた精子と産仔およびその子孫
- 左上:マウス多能性幹細胞(ES細胞)から試験管内で誘導した精子幹細胞様細胞(GSCLCs)は、精子へと分化する能力を保持することが示された。
- 左下:ES細胞由来である証拠に精子頭部が緑色に光っている。
- 右上:さらに、得られた精子は顕微授精により健常な産仔となることが示された。
- 右下:またその産仔は成体へと成長後、健常な子孫を産出することができた。
<用語解説>
- 注1) 多能性幹細胞
- 自己複製能力と、身体を構成するほぼ全ての細胞に分化する能力を持つ細胞のこと。胚性幹細胞(Embryonic stem cells:ESCs)や人工多能性幹細胞(induced Pluripotent stem cells:iPSCs)の総称。
- 注2) 精子幹細胞
- 自己複製能力と精子分化能力を併せ持った、オスの生涯にわたる精子産生の元となる細胞。精子幹細胞は精原細胞、精母細胞を経て、精子細胞へと分化する。
- 注3) 精子幹細胞様細胞
- 本研究において、マウス多能性幹細胞(ES細胞)から完全な試験管内で誘導した、精子幹細胞によく似た性質を持つ細胞。この精子幹細胞様細胞の長期培養株はGermline stem cell-like cells(GSCLCs)と呼ばれ、マウスの生後7日齢の精子幹細胞から樹立したGS細胞に近い細胞であることが示された。
- 注4) 始原生殖細胞
- 精子や卵子の元であり、生殖細胞系列の起点となる細胞。マウスでは、受精後6.25日後頃にエピブラスト注13)から出現する。
- 注5) 始原生殖細胞様細胞
- Primordial germ cell-like cells(PGCLCs)と呼ばれる。多能性幹細胞から、完全な試験管内で誘導した、始原生殖細胞に非常によく似た性質を持つ細胞。マウスの受精後8.5~9.5日齢の始原生殖細胞に相同であることが、転写産物およびエピゲノム状態の解析から示されている。
- 注6) 生殖巣
- 生殖細胞とそれらを支持する体細胞からなる構造体。母胎で、胎仔の始原生殖細胞におけるオス、メスの性分化が始まる頃(マウスでは受精後12.5日齢)までに形成される。始原生殖細胞はオス、メス各々に特徴的な生殖巣の体細胞に包まれながら、配偶子(精子や卵子)へと分化する。
- 注7) DNAメチル化
- エピゲノム情報の一つ。DNAの塩基配列を構成する4つの塩基(アデニン、シトシン、グアニン、チミン)のうち、シトシンの5位の炭素がメチル化されること。一般的には、このメチル化修飾に結合するタンパク質の働きなどにより、遺伝子の発現が抑制される。
- 注8) エピゲノム情報の再構成
- ゲノム(DNAの塩基配列)に付帯する、修飾情報の消去および再獲得のこと。修飾情報の代表として、DNAのメチル化やヒストンの修飾がある。
- 注9) GS細胞(Germline stem cells)
- 生体由来の精子幹細胞そのものを、長期間培養しうる細胞株にしたもの。凍結保存が可能で、遺伝情報を保ったまま2年以上安定的に増殖させることができる。これまで、マウス、ラット、ハムスター、ウサギにて培養株の樹立が報告されている。
- 注10) 前精原細胞
- オスの胎仔生殖巣内で精子幹細胞への分化を開始した生殖細胞のこと。始原生殖細胞が胎仔精巣体細胞に囲まれると、前精原細胞となり、生後になると精子幹細胞や精原細胞へと分化する。
- 注11) 精原細胞
- オスの生後における精巣内の生殖細胞のうち、精子幹細胞以外の未分化な細胞のこと。精原細胞は精巣体細胞に支持されながら精子へと分化する。
- 注12) 転写産物
- 設計図であるゲノム(DNA配列)と、実際に生体内で働くタンパク質とをつなぐ、中間産物。主にメッセンジャーRNAを指す。
- 注13) エピブラスト
- 内部細胞塊から分化した、体を構成する三胚葉(外胚葉、中胚葉、内胚葉)全てに分化する能力を持つ細胞集団。
<論文情報>
| タイトル |
“In Vitro Derivation and Propagation of Spermatogonial Stem Cell Activity from Mouse Pluripotent Stem Cells.” |
| 著者名 |
石藏 友紀子、薮田 幸宏、大田 浩、林 克彦、中村 友紀、岡本 郁弘、山本 拓也、栗本 一基、白根 健次郎、佐々木 裕之、斎藤 通紀 |
| doi |
10.1016/j.celrep.2016.11.026 |
<お問い合わせ先>
<研究に関すること>
斎藤 通紀(サイトウ ミチノリ)
京都大学 大学院医学研究科 生体構造医学講座(機能微細形態学) 教授
Tel:075-753-4335(直通) Fax:075-751-7286
E-mail:
<JST事業に関すること>
大山 健志(オオヤマ タケシ)
科学技術振興機構 研究プロジェクト推進部
Tel:03-3512-3528 Fax:03-3222-2068
E-mail:
<報道担当>
京都大学 企画・情報部広報課 国際広報室
Tel:075-753-5727 Fax:075-753-2094
E-mail:
科学技術振興機構 広報課
Tel:03-5214-8404 Fax:03-5214-8432
E-mail: