ポイント
- 植物の受精卵が分裂し成長する様子をリアルタイムで観察することに初めて成功した。
- 胚がダメージを受けたときに細胞運命を変えて植物が再生する能力を明らかにした。
- 高い再生能力を生み出す仕組みの解明、育種・培養技術の開発への貢献が期待される。
JST 戦略的創造研究推進事業において、名古屋大学 WPIトランスフォーマティブ生命分子研究所の東山 哲也 教授と名古屋大学 大学院理学研究科の栗原 大輔 特任助教らは、植物の受精卵が分裂し発生する様子を生きたままリアルタイムで観察(ライブイメージング)できるシステムを開発しました。
多細胞生物の複雑な器官は、受精卵という単細胞に由来します。動物の受精卵は取り出すことが容易なため、分裂していく過程(胚発生注1)過程)を生きたまま詳細に観察できます。しかし、被子植物の受精卵は母体組織であるめしべの奥深くに埋め込まれているので、受精卵の分裂過程を生きたまま観察することができません。このため、植物胚発生研究の歴史は長いにもかかわらず、実際にどのように細胞が分裂し、胚発生が進行しているのか、詳細は明らかではありませんでした。
研究グループは、植物の成長活性を損なわない培地の開発、高度な顕微鏡法、そしてマイクロ加工技術による新たなマイクロデバイス(マイクロピラーアレイ注2))を開発し、将来種子になる組織の中で、受精卵が分裂し胚を形成していく様子を生きたまま映像として捉えることに世界で初めて成功しました。また、レーザー技術でごく初期の胚を破壊すると、胚の隣に存在する細胞が発生運命を転換注3)し、再び胚を形成する驚くべき再生能力を観察することにも成功しました。
本研究成果は、植物胚発生や細胞運命転換の研究における基盤技術となり、植物の優れた再生能力の仕組みの解明や、育種・培養技術の開発につながると期待されます。
本研究成果は、2015年7月9日(米国東部時間)に米科学誌「Developmental Cell」のオンライン速報版で公開されます。
本成果は、以下の事業・研究領域・研究課題によって得られました。
戦略的創造研究推進事業 総括実施型研究(ERATO)
| 研究プロジェクト |
「東山ライブホロニクスプロジェクト」 |
| 研究総括 |
東山 哲也(名古屋大学 WPIトランスフォーマティブ生命分子研究所 教授) |
| 研究期間 |
平成22年10月~平成28年3月 |
多細胞生物の細胞が種々のシグナル分子を介して、他の細胞と情報のやり取りをすることで自身の振る舞いを決定し、生物としてのバランスを取るための仕組み「ホロニックコミュニケーション」の全容解明に向けた、多くのライブセル解析技術の創出を目指します。
<研究の背景と経緯>
多細胞生物には複雑な構造や機能を持つさまざまな器官がありますが、それらはすべて受精卵というたった1つの細胞に由来しています。受精卵が細胞分裂を繰り返し、からだを作っていく過程を胚発生過程と言います。被子植物において、受精卵の不等分裂により生じる2つの細胞、頂端細胞注4)と基部細胞注5)は形態的特徴も異なりますが、その後の発生運命も大きく異なります。頂端細胞は、分裂面を高頻度に変化させながら特有の細胞パターンを示す胚を形成し、植物体を生み出します。すなわち、頂端細胞は胚始原細胞と言えます。一方、基部細胞は一方向にのみ数回分裂し、母体組織とつながる胚柄を形成し胚への栄養供給を担います(図1)。ごく初期であるわずか2細胞の時期において、異なる細胞運命を持つようになりますが、その細胞運命を決定する仕組みについては、不明な点が多く残されています。近年、個々の細胞は他の細胞とコミュニケーションを取りながら、それぞれの細胞運命を決定し、からだ作りをすることが示唆されていますが、その詳細はまだ明らかになっていません。このように、胚発生過程は生命の根幹をなしますが、植物の受精卵は花の奥深くに埋め込まれているため、動物のように受精卵の分裂から胚発生までの一連の過程をリアルタイムで観察することはできませんでした。
<研究の内容>
研究グループは、モデル植物であるシロイヌナズナを用いて、将来種子となる母体組織(胚珠注6))をめしべから取り出して、胚珠の中で胚発生を再現することを試みました。まず、胚発生に異常なく胚珠が成長できる培地成分を検討しました。胚珠を包んでいる組織である子房注7)の培養に用いられてきた培地に、有機物としてトレハロースを加えたところ、胚珠生存率および正常な胚発生率の劇的な上昇が観察されました。この新しい胚珠培養培地の開発によって、めしべから取り出した胚珠内での胚の成熟および発芽、また発芽した個体を成熟個体まで栽培することも可能となりました。
植物の胚発生は数日に渡る過程であるために、長時間安定して顕微鏡観察する技術が必要でしたが、胚珠は楕円形をしているため観察中に視野からずれてしまう問題点がありました。研究グループは、これまでにマイクロ加工技術により、胚珠を安定して保持できるマイクロケージアレイ注8)を作製していました。本研究では改良を加えたマイクロピラーアレイを開発することによって、胚珠の成長を妨げることなく、長時間安定して胚珠を保持、培養する系を確立しました(図2)。マイクロピラーアレイはマイクロケージアレイよりも柔軟な構造のため、胚珠への影響をより少なく、また効率よく数多くの胚珠を保持することが可能となりました。
このように確立した胚珠培養技術と、何層もの細胞により厚く覆われた胚珠の中にある胚を高感度に撮影できる顕微鏡システムを組み合わせることによって、受精卵から後期胚までを通じて胚発生のライブイメージングに初めて成功しました(図3)。
またライブイメージングによって個々の細胞の分裂周期を解析し、シロイヌナズナ初期胚発生の細胞分裂系譜注9)を作製しました(図4)。受精卵の不等分裂で生じた頂端細胞は10.7時間後、もう一方の基部細胞は12.8時間後に再び分裂しました。分裂するまでの時間が異なることからも頂端細胞と基部細胞は受精卵分裂直後から異なる細胞特性を持っていることが分かりました。これまでシロイヌナズナの初期胚細胞は、それぞれ同じ周期で細胞分裂していると考えられてきましたが、頂端細胞の4回目の分裂からそれぞれの細胞の位置に応じて分裂周期が異なり始める様子が観察され、ごく初期からそれぞれの細胞特性を獲得していることが分かりました。
受精卵の不等分裂により生じた頂端細胞と基部細胞はすでに異なる細胞運命を獲得していますが、それらの細胞運命は一度決まってしまったら変わらないのか、それとも変わりうるのかを明らかにするために、受精卵が分裂した直後の細胞1つのみを顕微鏡下でレーザーによって破壊し、解析しました。受精卵は厚い胚珠組織に覆われているため、研究グループは組織深部にも照射できるフェムト秒パルスレーザー注10)という特殊なレーザーを用いて、受精卵が分裂した直後の頂端細胞を破壊し、3日間に渡り、その後の影響をタイムラプス観察(連続観察)しました。
頂端細胞をレーザーで破壊し、残った基部細胞をタイムラプス観察した結果、残った基部細胞はまず胚柄細胞特有の分裂をした後、驚くべきことに上側の胚柄細胞が胚細胞特有の細胞分裂様式を示しました。この細胞の特性を調べるために、受精卵分裂後、胚柄細胞でのみ発現し胚柄細胞の遺伝子発現を制御する転写因子WOX8 遺伝子、そして胚細胞でのみ発現する転写因子DRN 遺伝子の発現を解析しました。頂端細胞を破壊した後すぐは、残った基部細胞はWOX8 遺伝子を発現したままでしたが、1回分裂した後に上側の細胞でWOX8 遺伝子の発現が低下し、代わりにDRN 遺伝子の発現が上昇、そして胚細胞特有の細胞分裂を行いました(図5)。このことにより、頂端細胞がダメージを受けると、隣にあるすでに胚柄細胞へと細胞運命が決定した細胞が、失った頂端細胞を補うために新たに頂端細胞へと運命転換が起こることが明らかとなりました。最終的に植物体を作り上げる頂端細胞でさえ、失われた場合には1細胞からでも再び作り直すことができる、植物の驚くべき再生能力を示した研究結果と言えます。
<今後の展開>
胚発生研究は植物科学分野で古くから世界中で研究されてきた分野の1つです。しかし、これまで胚を生かしたまま解析することがかなわなかったため、どのように細胞同士がコミュニケーションを取りながら植物体を作り上げるのか明らかではありませんでした。今回開発した植物受精卵・胚発生のライブイメージング・光顕微操作システムを用いることで、世界中で胚発生研究が加速していくと期待されます。
本研究成果により、被子植物において1細胞からでも植物体を作り直すことができる再生能力の存在が明らかとなりました。今後は、細胞の運命転換を誘導し再生する分子メカニズムを明らかにしていくことが焦点になります。農業や園芸の分野で古くから用いられている組織培養からも明らかなように、植物には高い再生能力が備わっていることが知られています。細胞の運命転換を可能にする分子メカニズムを明らかにすることで、効率の良い組織培養など、植物にかかわるさまざまな育種・培養技術開発への貢献も期待されます。
<参考図>
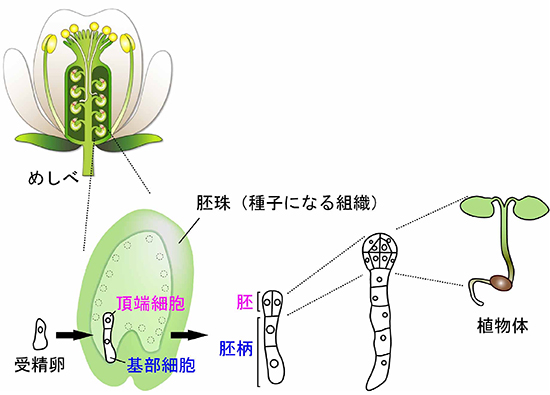
図1 シロイヌナズナの花と胚発生の模式図
被子植物において、将来種子になる胚珠の中で胚発生は進行します。受精卵は著しく細胞伸長した後、小さい頂端細胞と大きい基部細胞へと大きさの異なる2つの細胞に分裂(不等分裂)します。頂端細胞は胚を形成し、最終的には植物体を形成します。
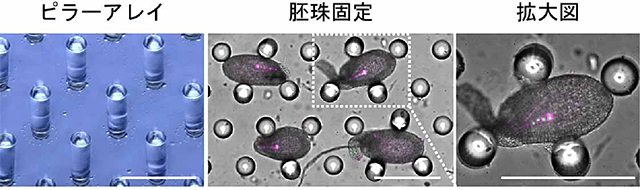
図2 マイクロデバイスによる胚珠固定
ピラーが等間隔に並んだマイクロピラーアレイを用いることによって、長時間安定して胚珠を保持することができます。ピラーは胚珠の成長を妨げることなく胚珠を保持するので、胚発生の詳細な観察が可能です。スケールバーは300マイクロメートルを表しています。
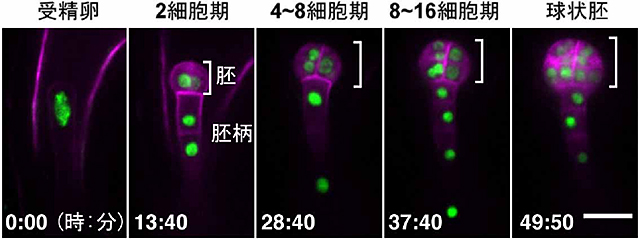
図3 受精卵分裂と胚発生のライブイメージング
受精卵は分裂し、胚と胚柄を形成します。緑色は細胞核を、ピンク色は細胞膜を示しています。胚細胞は分裂する方向を変えながら丸い組織を作っていきますが、胚柄細胞は縦にのみ分裂し、棒状の組織を作っていきます。胚に2つ細胞があるときに2細胞期と呼びます。数字は観察開始からの時間、スケールバーは20マイクロメートルを表しています。
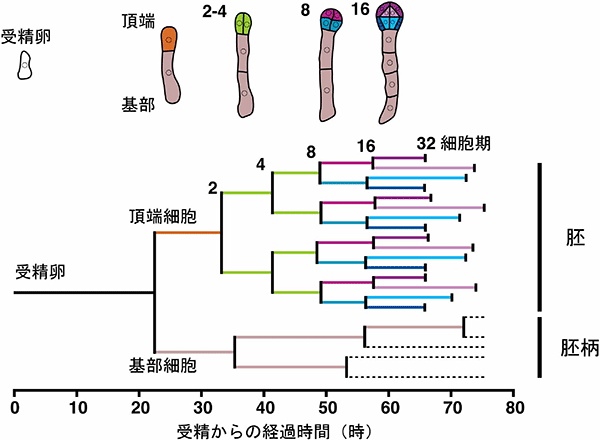
図4 受精からの細胞分裂系譜図
横軸の長さはそれぞれの細胞が分裂にかかる時間を示しています。バーの色は上図の細胞の位置を示しています。点線部分は未計測です。頂端細胞から発生する胚細胞は、2−4細胞期ではそれぞれ同じ周期(緑)で分裂していますが、8細胞期では上(紫)と下(青)、16細胞期では内(薄い色)と外(濃い色)で分裂周期が異なっていることが分かります。また、基部細胞から発生する胚柄細胞の分裂周期は胚細胞とずれていることも分かります。
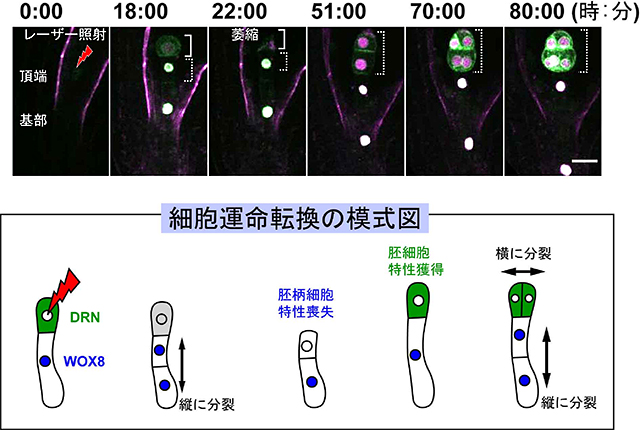
図5 頂端細胞破壊により誘導される細胞運命転換
フェムト秒パルスレーザー照射により頂端細胞を破壊しました。写真の実線枠はレーザー照射した胚を示します。22時間後、細胞は完全に萎縮しました。点線枠で示している細胞では、胚柄マーカーWOX8(白、模式図では青:核に存在)の発現が減少し、胚マーカーであるDRN(緑:細胞質に存在)の発現が上昇し、基部細胞由来にもかかわらず胚細胞特有の分裂を示しました。数字はレーザー照射後からの経過時間、スケールバーは20マイクロメートルを表しています。
<用語解説>
- 注1) 胚発生
- 被子植物においては、種子の中で受精卵から幼植物が作られる過程です。
- 注2) マイクロピラーアレイ
- 微小な円柱(ピラー)が並んでいるデバイスです。複数の円柱で組織を取り囲むことにより安定して保持することが可能です。また円柱は柔軟な素材でできているため、胚珠の成長を妨げません。
- 注3) 細胞運命転換
- 特定の細胞種へと分化する運命(細胞運命)が、別の細胞種へ分化する運命に変わることです。
- 注4) 頂端細胞
- 受精卵が不等分裂してできる細胞質に富んだ小さい細胞です。活発に細胞分裂を繰り返して胚を形成し、最終的に植物体を形成します。
- 注5) 基部細胞
- 受精卵が不等分裂してできる液胞に富んだ大きな細胞です。一方向に数回分裂して棒状の胚柄を形成します。胚柄は胚と母体組織である胚珠をつなぐ役割を持っており栄養供給を担いますが、最終的には退化して消失します。
- 注6) 胚珠
- めしべの奥深くに位置する種子のもととなる組織です。胚珠の中にある卵細胞と花粉管によって運ばれてきた精細胞が受精することによって、受精卵が作られます。その後、受精卵が分裂を繰り返し胚発生が進行します。
- 注7) 子房
- めしべの下部の膨らんだ組織で、成長すると果実になります。子房の中に胚珠があり、胚珠は成長すると種子になります。
- 注8) マイクロケージアレイ
- 微小な檻(ケージ)型の構造物が並んでいるデバイスです。胚珠を檻の中に閉じこめることにより、安定して保持することができます。
- 注9) 細胞分裂系譜
- 受精卵分裂から始まる胚発生の過程で、各細胞がどのタイミングで分裂し、どの細胞になるのかを記述したものです。
- 注10) フェムト秒パルスレーザー
- フェムト秒というごく短いパルス幅で発振されるレーザーであるため、瞬間的に高い光強度を持ちます。また長波長の近赤外光を用いることから、生体内における散乱の影響を受けにくく、組織深部までレーザーを照射することが可能です。
<論文タイトル>
“Live-Cell Imaging and Optical Manipulation of Arabidopsis Early Embryogenesis”
(シロイヌナズナ初期胚発生過程のライブセルイメージングと光顕微操作)
doi :10.1016/j.devcel.2015.06.008
<お問い合わせ先>
<研究に関すること>
東山 哲也(ヒガシヤマ テツヤ)
名古屋大学 ERATO 東山ライブホロニクスプロジェクト 研究総括
名古屋大学 WPIトランスフォーマティブ生命分子研究所 教授
〒464-8602 愛知県名古屋市千種区不老町
Tel:052-747-6404 Fax:052-789-2497
E-mail:
栗原 大輔(クリハラ ダイスケ)
名古屋大学 大学院理学研究科 特任助教
名古屋大学 ERATO 東山ライブホロニクスプロジェクト グループリーダー
Tel:052-747-6401 Fax:052-789-2497
E-mail:
<JST事業に関すること>
大山 健志(オオヤマ タケシ)
科学技術振興機構 研究プロジェクト推進部
〒102-0076 東京都千代田区五番町7 K’s五番町
Tel:03-3512-3528 Fax:03-3222-2068
E-mail:
<報道担当>
科学技術振興機構 広報課
〒102-8666 東京都千代田区四番町5番地3
Tel:03-5214-8404 Fax:03-5214-8432
E-mail:
名古屋大学 総務部 広報渉外課
〒464-8601 愛知県名古屋市千種区不老町
Tel:052-789-2699 Fax:052-788-6272
E-mail: