理化学研究所(理研) 光量子工学研究領域ライブセル分子イメージング研究チームの岩井 優和 客員研究員(JSTさきがけ研究者)、中野 明彦 チームリーダーらの共同研究グループ※は、コケ植物の光合成反応を担う光化学系タンパク質注1)の解析を行い、コケ植物特有の「集光アンテナ調節機構」を解明しました。
光合成反応は、植物の葉緑体で行われる化学反応で、自然環境の維持と物質生産という重要な役割を担っています。光合成反応に関わる光化学系Ⅰと光化学系Ⅱ注2)は、葉緑体のチラコイド膜注3)に存在するタンパク質複合体で、集光アンテナタンパク質注4)から運ばれた光エネルギーを消費して光合成電子伝達系を駆動させ、エネルギー生産を行います。それぞれの光化学系の反応効率を高めるため、集光アンテナタンパク質が光エネルギーの供給量を調節しています。これを「集光アンテナ調節機構」と呼びます。これまで、緑藻と陸上植物の集光アンテナタンパク質について盛んに研究が進められてきましたが、進化的に陸上植物と緑藻の中間に位置するコケ植物の集光アンテナタンパク質については研究が進んでいませんでした。
共同研究グループは、進化的に緑藻と陸上植物の間に位置するコケ植物「ヒメツリガネゴケ注5)」のチラコイド膜を分離精製し、解析を行いました。その結果、ヒメツリガネゴケのチラコイド膜には、陸上植物型と緑藻型の2種類の光化学系Ⅰ複合体が存在することが分かりました。また、ヒメツリガネゴケにのみ存在が確認されているLhcb9と呼ばれる集光アンテナタンパク質が、緑藻型光化学系Ⅰ複合体の形成に重要であることも分かりました。ゲノム情報が公開されている全ての光合成真核生物注6)の集光アンテナタンパク質との進化的な関係を調べたところ、ヒメツリガネゴケが持つLhcb9は、その元となる遺伝子を自らの祖先ではなく、緑藻の祖先から水平伝播注7)によって獲得していたことが示唆されました。そのため、ヒメツリガネゴケは緑藻型光化学系Ⅰ複合体を持っていると考えられます。陸上植物型と緑藻型の2つの光化学系Ⅰ複合体を持つことの進化的な意義は不明ですが、植物が水中から陸上に進出する際の環境変化の中で、2つの光化学系Ⅰ複合体を持つことで電子伝達系によるエネルギー生産を助けていた可能性があります。今回の研究成果から、進化の過程で獲得した新規の集光アンテナタンパク質が光合成反応の活性に影響を及ぼすことが分かりました。
本研究は、科学研究費補助金(若手研究(A):課題番号23687008)の支援を受けて行われ、JST 戦略的創造研究推進事業 個別型研究(さきがけ)研究領域「二酸化炭素資源化を目指した植物の物質生産力強化と生産物活用のための基盤技術の創出」研究課題名「ライブセルイメージングによる光環境適応機構の実態解明」(代表:岩井 優和)の一環として行われました。成果は、英国の科学雑誌「Nature Plants」のオンライン版で1月19日(現地時間)に掲載されます。
<背景>
約30億年という長い年月の間、植物などの光合成生物は地球規模で起こるさまざまな環境変化を耐え抜き、あらゆる自然環境下で生存し続けてきました。光合成生物がどのように環境の変化に適応してきたのかを調べることで、今後、予測される環境変化にも耐えうる植物の開発を進めることができます。多くの植物は、さまざまな形態形成によって環境への適応を図ってきましたが、光合成反応を司る葉緑体タンパク質も環境に合わせて、その機能を調節していたことが知られています。これまで研究されてきたほとんど全ての光合成生物では、光合成反応を司る根幹部分はよく似ているのですが、光エネルギーを吸収した後の処理の仕方が異なっていることが、近年明らかになっています。
光エネルギーを吸収するためのクロロフィル(葉緑素)を多く含み、吸収した光エネルギーを、葉緑体のチラコイド膜に存在する「光化学系」へと運ぶ重要な役割を担うのが「集光アンテナ(LHC)タンパク質」です。光化学系は、集光アンテナタンパク質から運ばれた光エネルギーを消費して光合成電子伝達系を駆動させ、エネルギー生産を行います(図1)。しかし、集光アンテナタンパク質から必要以上に光エネルギーが供給されると光化学系は活性酸素によって壊れてしまいます。それを防ぐため、集光アンテナタンパク質は過剰な光エネルギーの行き先をコントロールして光化学系の機能を維持しています。これを「集光アンテナ調節機構」と呼びます。
光化学系には、光化学系Ⅰと光化学系Ⅱの2種類があり、それぞれに光エネルギーを供給する集光アンテナタンパク質をLHCⅠ、LHCⅡ注8)と呼んでいます(図1)。LHCⅡは、地球上で最も多い膜タンパク質として知られ、光エネルギーの制御に重要な役割を持っていることがこれまでの研究で分かっています。また、LHCⅡの基本骨格を持つ集光アンテナタンパク質はほとんどの光合成真核生物に保存されていますが、進化の過程で多様化しています。この集光アンテナタンパク質の多様性が、さまざまな環境への適応を可能としているのではないかとも考えられています。
これまで緑藻や陸上植物の集光アンテナタンパク質について盛んに研究されてきましたが、進化的に緑藻と陸上植物の中間に位置するコケ植物の同タンパク質についてはほとんど研究が進んでいません。そこで共同研究グループは、コケ植物特有の環境適応機構の解明を目的に、ヒメツリガネゴケの集光アンテナタンパク質の解析を行いました。
<研究手法と成果>
光合成反応の光エネルギー伝達に関わるタンパク質のほとんどは葉緑体のチラコイド膜に存在しています。共同研究グル-プはまず、ヒメツリガネゴケのチラコイド膜の光合成関連タンパク質をショ糖密度勾配超遠心法注9)によって分離精製しました。その結果、ヒメツリガネゴケには分子量の大きさが異なる2つの光化学系Ⅰ複合体が存在していることが分かりました。
この2つの光化学系Ⅰ複合体に含まれる集光アンテナタンパク質を調べたところ、分子量の大きい光化学系Ⅰ複合体には、光化学系Ⅰひとつに対して多くのLHCⅠが含まれていることが分かりました。これは、緑藻の光化学系Ⅰ複合体の特徴と一致しています。また、多くのLHCⅠが含まれるだけでなく、ヒメツリガネゴケ特有のLHCⅡであるLhcb9も含まれていることが分かりました。そこで、Lhcb9を欠損させた変異株を作成し、同様に解析した結果、Lhcb9欠損株では分子量の大きい光化学系Ⅰ複合体がなくなり、分子量の小さい光化学系Ⅰ複合体しか存在していませんでした。このことから、分子量の大きい光化学系Ⅰ複合体の形成には、Lhcb9が重要であることが分かりました。
次に、Lhcb9と分子量の大きい光化学系Ⅰ複合体が物理的に結合しているかを調べるため、Lhcb9に目的のタンパク質を分離できるヒスチジンタグ注10)を付けた変異株を作成しました。ヒスチジンとの親和性の高いニッケルが吸着した樹脂を用いて、Lhcb9を単離精製したところ、光化学系Ⅰ複合体も一緒に単離されたことから、Lhcb9と光化学系Ⅰ複合体は物理的に結合していることが分かりました。また、Lhcb9が吸収した光エネルギーが光化学系Ⅰ複合体へ伝達されているかを調べるため、分光学的解析を行いました。その結果、Lhcb9が吸収した光エネルギーは効率良く光化学系Ⅰ複合体へ伝達されていることが分かりました。また、過剰な光エネルギーを吸収した際に、Lhcb9は分子量の大きい光化学系Ⅰ複合体から他の集光アンテナタンパク質を分離している役割をもっていることも分かりました。これらのことから、Lhcb9は光化学系Ⅰ複合体の光エネルギー吸収量を高めるだけでなく、光エネルギーの伝達量の調節も行っていることが分かりました(図2)。
さらに、ゲノム情報が公開されている全ての光合成真核生物の集光アンテナタンパク質とLhcb9の進化的な関係を系統樹解析注11)によって調べました。その結果、Lhcb9以外のヒメツリガネゴケが持つすべての集光アンテナタンパク質は、陸上植物のシロイヌナズナと最も近い構造をしていることが分かりました。しかし、Lhcb9だけは緑藻のクラミドモナスが持つLhcbm5と呼ばれるLHCⅡと最も近い構造でした。さらに、ヒメツリガネゴケはLhcb9の元となる遺伝子を、コケ植物の祖先からではなく緑藻の祖先から水平伝播によって獲得したことも示唆されました。
これまでの研究によって、緑藻型の光化学系Ⅰ複合体は、陸上植物型の光化学系Ⅰ複合体と比較して、LHCⅠが多数結合していることが分かっています。今回、ヒメツリガネゴケで単離に成功した分子量の大きい光化学系Ⅰ複合体にもLHCⅠが多数結合していました。詳細に解析した結果、ヒメツリガネゴケが持つ分子量の大きい光化学系Ⅰ複合体と、緑藻の光化学系Ⅰ複合体の分子量は同じで、分子量の小さい光化学系Ⅰ複合体と陸上植物の光化学系Ⅰ複合体の分子量が同じであることが分かりました(図3)。
緑藻が持つLhcbm5と最も相同性が高いLhcb9を欠損させると分子量の大きい光化学系Ⅰ複合体が形成できなくなるため、ヒメツリガネゴケが持つ分子量の異なる2つの光化学系Ⅰ複合体は、陸上植物型と緑藻型の光化学系Ⅰ複合体であることが示唆されました。
陸上植物型と緑藻型の2つの光化学系Ⅰ複合体を持つことが、ヒメツリガネゴケにとってどのような意義があるかは分かっていません。コケ植物の祖先から引き継がれず、水平伝播によって緑藻の祖先から獲得したLhcb9が緑藻型光化学系Ⅰ複合体を形成することから、コケ植物にとって緑藻と近い環境に適した光合成能力の獲得につながっている可能性があります。コケ植物が最初に水中から陸上に進出したと考えられていますが、大気と水中との2つ環境の狭間で生息していたことも想像できます。このことから、陸上植物型と緑藻型の2つの光化学系Ⅰ複合体を持つことで、激変する環境にその都度対応する必要がなかったのかもしれません。これまでの研究では、水平伝播による外来遺伝子の獲得がヒメツリガネゴケにとって陸上進出のきっかけになっている例が複数報告されていますが、集光アンテナタンパク質の遺伝子の水平伝播による獲得についての報告はこれが初めてです(図4)。また、2つの光化学系Ⅰ複合体を持つことは、ヒメツリガネゴケの集光アンテナ機構が陸上植物と緑藻とは異なることも示唆しています。従って、さまざまな環境に適応した光合成生物の集光アンテナ機構をより多く調べることで、まだ明らかになっていない光エネルギー制御機構が発見されるかもしれません。
<今後の期待>
光合成反応の根幹である光化学系の基本構造は、全ての光合成生物で保存されています。しかし光エネルギーを吸収し、光化学系に伝達する重要な役割を担う集光アンテナタンパク質の種類は多岐に富んでいます。これまでの研究で、陸上植物と緑藻の集光アンテナ調節については異なる機構が存在していることが知られていました。今回、ヒメツリガネゴケの集光アンテナ調節機構もコケ植物特有のものであることが分かりました。今後、さまざまな環境変化に耐え得る植物の作製を模索していく上で、多様な光合成生物の集光アンテナ調節機構を研究する必要があります。モデルにはまらない調節機構を持つ光合成生物が自然界にはまだ多く存在していることも十分に考えられます。今回のタンパク質解析はそれらを解明するための重要なアプローチの1つであり、これによって得られる情報が、生きた細胞内での光合成機能の実態解明の大きな手がかりとなります。
※ 共同研究グループ
理化学研究所 光量子工学研究領域
ライブセル分子イメージング研究チーム |
|---|
|
客員研究員 |
岩井 優和(いわい まさかず)(JSTさきがけ研究者) |
|---|
| チームリーダー |
中野 明彦(なかの あきひこ) |
| 北海道大学 低温科学研究所 |
|---|
|
研究員 |
横野 牧生(よこの まきお) |
|---|
| 神戸大学 分子フォトサイエンス研究センター |
|---|
|
准教授 |
秋本 誠志(あきもと せいじ) |
|---|
| 東京大学 大学院理学系研究科 生物科学専攻 植物生態学研究室 |
|---|
|
研究員 |
河野 優(こうの まさる) |
|---|
| 准教授 |
野口 航(のぐち こう) |
<参考図>
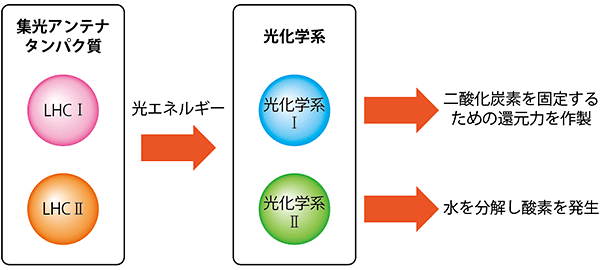
図1 集光アンテナタンパク質と光化学系
集光アンテナタンパク質は、吸収した光エネルギーを光化学系へ伝達する。光化学系Ⅰ・Ⅱは、その光エネルギーを消費して光合成電子伝達系を駆動させ、エネルギー生産を行う。
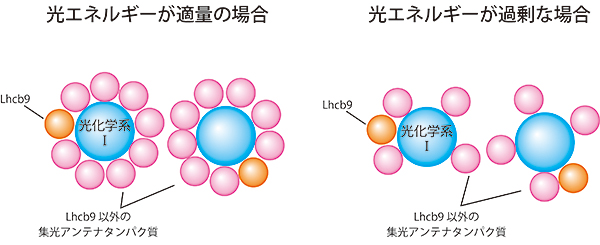
図2 ヒメツリガネゴケの光化学系Ⅰ複合体の集光アンテナ調節機構
ヒメツリガネゴケ特有のLHCⅡであるLhcb9は分子量の大きい光化学系Ⅰ複合体と物理的に結合している。Lhcb9が吸収した光エネルギーは効率良く光化学系Ⅰ複合体へ伝達される(左)。また、Lhcb9は分子量の大きい光化学系Ⅰ複合体が過剰な光エネルギーを吸収した際に、他の集光アンテナタンパク質を分離し、光エネルギーの伝達量を調節している。

図3 チラコイド膜に含まれる光化学系タンパク質の分離
シロイヌナズナ(陸上植物)、クラミドモナス(緑藻)、ヒメツリガネゴケ(コケ植物)のチラコイド膜に含まれる光化学系タンパク質などをショ糖密度勾配超遠心法により分離精製した。シロイヌナズナには分子量が小さい光化学系Ⅰ複合体のみ(矢印)が存在し、クラミドモナスには分子量が大きい光化学系Ⅰ複合体のみ(矢じり)が存在しているが、ヒメツリガネゴケには、2つとも存在していることが分かる。
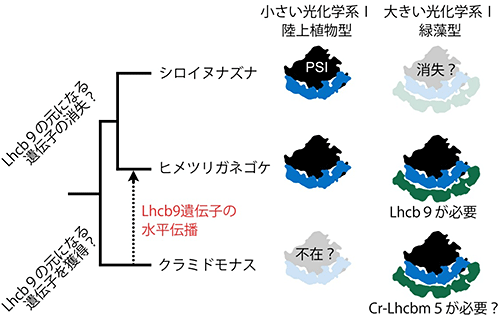
図4 光化学系Ⅰ複合体とLhcb9との進化的相関図
シロイヌナズナ、ヒメツリガネゴケ、クラミドモナスはそれぞれ元をたどれば、1つの共通の祖先から進化してきたことを前提にしている。最初の分岐点で、Lhcb9の元になる遺伝子を失うか、獲得するかで2つに枝分かれする。Lhcb9の元になる遺伝子を失った系統には、シロイヌナズナとヒメツリガネゴケが新たに分岐している。シロイヌナズナは分子量が小さい光化学系Ⅰ複合体のみ存在している。Lhcb9の元になる遺伝子を獲得した系統には、クラミドモナスが進化し、分子量が大きい光化学系Ⅰ複合体のみ存在している。進化の過程で、ヒメツリガネゴケはLhcb9遺伝子を緑藻の系統から水平伝播によって獲得した。その結果、ヒメツリガネゴケは分子量が小さい光化学系Ⅰと大きい光化学系Ⅰの両方を持つのではないかという仮説を示した。
<用語解説>
- 注1) 光化学系タンパク質
- 光エネルギーを使って化学反応を引き起こす酵素で、光合成反応において最も重要なタンパク質。
- 注2) 光化学系Ⅰと光化学系Ⅱ
- 光化学系Ⅰは、光エネルギーを利用して、二酸化炭素を固定するために必要な還元力を作る複数のサブユニットからなる光化学系タンパク質の1つ。光化学系Ⅱは、光エネルギーを利用して、水を分解し酸素を発生する複数のサブユニットからなる光化学系タンパク質の1つ。
- 注3) チラコイド膜
- 葉緑体内部にある脂質二重膜。光合成が起きる反応の場。
- 注4) 集光アンテナタンパク質
- クロロフィルを含む色素が結合したタンパク質複合体。光を吸収するアンテナとして働き、吸収した光エネルギーを光化学系タンパク質に伝達する。
- 注5) ヒメツリガネゴケ
- 進化的に陸上植物と緑藻の間に位置するコケ植物の1つ。コケ植物のモデル生物として初めてゲノムが解読され、2008年にゲノム情報が公開された。
- 注6) 光合成真核生物
- 光合成反応を行う真核生物。種子植物、シダ植物、コケ植物、シャジク藻、緑藻、褐藻(かっそう)、珪藻(けいそう)、渦鞭毛藻(うずべんもうそう)、クリプト藻、紅藻(こうそう)など。
- 注7) 水平伝播
- 親から子へ遺伝子が引き継がれるのではなく、異なる生物間で起こる遺伝子の移動・取り込みのこと。水平伝播によって獲得した遺伝子は、系統樹解析において整合性が取れないという特徴がある。
- 注8) LHCⅠとLHCⅡ
- LHCⅠは、光化学系Ⅰに主に結合し、光エネルギーを伝達する集光アンテナタンパク質の総称。LHCⅡは、光化学系Ⅱに主に結合し、光エネルギーを伝達する集光アンテナタンパク質の総称。LHCⅡは、チラコイド膜に存在するタンパク質で最も多く、地球上で最も多い膜タンパク質としても知られている。また、LHCⅡは過剰な光エネルギー下では、そのエネルギーを熱に変換する役割を持つことも知られている。
- 注9) ショ糖密度勾配超遠心法
- ショ糖の濃度が高いほうから低い順に遠心管に注ぎ、勾配を持つショ糖溶液を準備し、その上にさまざまな細胞内のタンパク質や高分子の物質を含む試料を載せ、高速で遠心することで、分子の大きさや沈降のしやすさなどの違いによって分離する方法。
- 注10) ヒスチジンタグ
- ヒスチジンはアミノ酸の一種。ヒスチジンタグは、ヒスチジンが6~8個ほど連なったペプチドで、遺伝子改変により目的のタンパク質の末端などに融合する。pHが8以上で、ニッケルなどの金属イオンが固定した樹脂(担体)と結合しやすいので、ヒスチジンタグを付けた目的のタンパク質を分離することができる。
- 注11) 系統樹解析
- アミノ酸配列や遺伝子配列、または生物が持つ特徴や形質に基づいて、生物間や遺伝子の進化的な道筋を推定する方法。樹状図を用いて、遺伝子や形質の差異から分岐点やその時期など推測する。
<論文情報>
“Light-harvesting complex Lhcb9 confers a green alga-type photosystem I supercomplex to the moss Physcomitrella patens”
Iwai, M., Yokono, M., Kono, M., Noguchi, K., Akimoto, S., Nakano, A.
Nature Plants
doi: 10.1038/NPLANTS.2014.8
<お問い合わせ先>
<研究に関すること>
岩井 優和(イワイ マサカズ)
理化学研究所 光量子工学研究領域 エクストリームフォトニクス研究グループ ライブセル分子イメージング研究チーム 客員研究員
中野 明彦(ナカノ アキヒコ)
理化学研究所 光量子工学研究領域 エクストリームフォトニクス研究グループ ライブセル分子イメージング研究チーム チームリーダー
Tel:048-467-9548 Fax:048-462-4679
E-mail:
<JST事業に関すること>
科学技術振興機構 戦略研究推進部
Tel:03-3512-3525 Fax:03-3222-2063
E-mail:
<報道担当>
理化学研究所 広報室 報道担当
Tel:048-467-9272 Fax:048-462-4715
E-mail:
科学技術振興機構 広報課
Tel:03-5214-8404 Fax:03-5214-8432
E-mail: