ポイント
- 受精を示す細胞内のカルシウム濃度の変化を、初めて被子植物で観察。
- 受精に関わる個々の細胞が協調して重複受精を達成する様子が明らかに。
- 受精の進行が明確になることで植物の受精や発生開始の仕組みの解明に期待。
JST 戦略的創造研究推進事業において、名古屋大学の東山 哲也 教授、浜村 有希 研究員(現 モントリオール大学)らは、植物の受精に伴う細胞内のカルシウム濃度変化を、種子になる組織の中で観察することに初めて成功しました。
カルシウムは細胞内の情報伝達の役割を担うことが知られています。動物では精細胞と融合するとき、つまり受精の瞬間の卵細胞でカルシウム濃度の上昇や、受精卵でのカルシウム振動(カルシウム濃度の周期的な変化)などが詳しく解析されてきました。このようなカルシウム濃度変化から、受精の進行を知ることができます。しかし植物では、受精に伴ってどのようなカルシウム濃度変化があるか解明されていませんでした。
被子植物の受精は、「重複受精」と呼ばれ、めしべの奥で進行します。花粉がめしべにつくと、花粉から花粉管という細い管が伸び、卵細胞の隣にある2つの助細胞に導かれて種子になる組織(胚珠)に向かいます。その管の中を2個の精細胞が移動して、1つが卵細胞と、もう1つは中央細胞と受精します。受精した卵細胞は、幼植物になる「胚」に、中央細胞は種子の発芽の際に養分を供給する「胚乳」になります。
研究チームは、胚珠を用いた高効率な体外重複受精と、高感度な顕微鏡を用いたカルシウム濃度観察法により、受精に伴うカルシウム濃度変化を観察することに成功しました。花粉管が到達すると2つの助細胞でカルシウム振動が始まり、一方で濃度が高まると花粉管から精細胞の放出が起こりました。この瞬間、卵細胞、中央細胞、2つの助細胞の全ての細胞でカルシウム濃度が最大値を示し、カルシウム濃度がより高まっていた方の助細胞は崩壊しました。続いて卵細胞では、受精に伴い一度だけカルシウム濃度の上昇が観察されました。受精によってカルシウム振動が始まる動物とは異なり、植物では一度だけの濃度上昇により、2つ目の精細胞との受精を防ぎ、胚発生開始のタイミングが制御されている可能性が考えられます。また、興味深いことに、残った助細胞で再びカルシウム振動が見られました。このことは2本目の花粉管の誘引の停止に関わる可能性が考えられます。これらの知見は、組織深部で進行するために、謎の多い植物の重複受精の仕組みの解明につながると期待されます。
本研究成果は、2014年8月22日(英国時間)に英科学誌「Nature Communications」のオンライン速報版で公開されます。
本成果は、以下の事業・研究プロジェクトによって得られました。
戦略的創造研究推進事業 総括実施型研究(ERATO)
| 研究プロジェクト |
「東山ライブホロニクスプロジェクト」 |
| 研究総括 |
東山 哲也(名古屋大学 トランスフォーマティブ生命分子研究所 教授) |
| 研究期間 |
平成22年~平成27年 |
上記研究課題では、ライブセル解析による多細胞生物のホロニックコミュニケーションの全容解明に挑みます。多細胞生物の細胞は、種々のシグナル分子を介して、ほかの細胞と情報のやりとりをすることで自身の振る舞いを決定し、生物としてのバランスを取っています。こうした個と全体を調和する細胞間コミュニケーションを、ホロニックコミュニケーションと呼びます。そのメカニズムの解明および解析技術の確立は、広く生物学や医療の分野での貢献が期待できます。本研究では、そのための多くのライブセル解析技術の創出を目指します。
<研究の背景と経緯>
被子植物の花粉は精細胞を2つ含み、花粉から伸びる花粉管により種子になる組織(胚珠)に運ばれます。花粉管は胚珠の中の卵細胞の隣に2つある助細胞により卵細胞の近くへ誘引されます。花粉管が卵細胞に到達すると、花粉管の先端が破裂して精細胞が放出されます。それと同時に、一方の助細胞が崩壊して受精の場が作られます。2つある精細胞のうち一方が卵細胞と受精して胚になり、もう一方は卵細胞の隣にある中央細胞と受精して胚乳を作る、被子植物の特徴的な受精は重複受精と呼ばれています(図1)。
細胞内のカルシウム濃度の変化は、細胞内でシグナル伝達が起こっていることを示す重要な指標です。動物においては、受精に伴う卵細胞のカルシウム濃度上昇や受精卵におけるカルシウム振動が見られることが詳しく解析されてきましたが、被子植物の重複受精に深く関わる卵細胞、中央細胞、2つの助細胞で、受精に伴ってどのようなカルシウム濃度の変化があるかは明らかでありませんでした。
研究チームは、これまでにもアブラナ科のシロイヌナズナで、受精に関わる細胞を種子となる組織から単離することなく重複受精を再現する、体外重複受精の技術を用い、受精時の精細胞の動きを観察していきました。この体外重複受精の技術と、カルシウム濃度を検出できる蛍光たんぱく質、さらに厚く覆われた胚珠内を高速かつ高感度に撮影できる顕微鏡システムを組み合わせることで、種子となる組織の中にある4つの細胞の、受精時のカルシウム濃度の変化を観察することが可能になりました。
<研究の内容>
卵細胞、中央細胞そして2つの助細胞でカルシウム濃度を検出する蛍光たんぱく質を働かせる植物をそれぞれ作り、まず、卵細胞のカルシウム濃度変化を観察しました。すると、卵細胞では2回のカルシウム濃度上昇があることが分かりました。このカルシウム濃度上昇のタイミングが重複受精のどの段階に関係しているか調べるために、精細胞の動きとカルシウム濃度変化を同時に観察しました。精細胞が大きく動く花粉管の精細胞放出と1回目のカルシウム濃度上昇のタイミングを比べると、その差は、2.3±15秒でほぼ一致していたので、この1回目のカルシウム濃度上昇は花粉管の精細胞放出に伴って起きていることが分かりました(図2)。
次に、2回目のカルシウム濃度上昇のタイミングと、卵細胞の受精、中央細胞の受精のタイミングを比べると、2回目のカルシウム濃度上昇は、卵細胞の受精と強い相関が見られました。そこで、2回目のカルシウム濃度上昇が卵細胞の受精に伴うものであることを調べるため、精細胞が1つしかできないため、卵細胞か中央細胞のいずれかにしか受精できない変異体(cdka;1)を用いて2回目のカルシウム濃度変化を観察しました。すると、cdka;1 変異体では、卵細胞と受精した時のみ2回目のカルシウム濃度上昇が検出されました。これによって、確かに、卵細胞の2回目のカルシウム濃度上昇は、卵細胞の受精に関係しているが、中央細胞の受精には関連がないことが分かりました。
さらに、卵細胞と精細胞の膜融合注1)と卵細胞のカルシウム濃度上昇の正確なタイミングを調べるため、卵細胞の膜とカルシウム濃度を検出する蛍光たんぱく質を組み合わせて観察したところ、2回目のカルシウム濃度上昇と、精細胞が卵細胞膜を横切るタイミングとが一致していました(図3)。これによって、卵細胞における2回目のカルシウム濃度上昇が、精細胞との膜融合によって起こることが明らかになりました。
このように、植物の卵細胞では、受精に伴いカルシウム振動が始まる動物とは異なるカルシウム濃度変化が見られました。この単発のカルシウム濃度上昇により、2つ目の精細胞との受精を防ぐこと(多精拒否)や発生開始のタイミングが制御されているのかもしれません。
次に、重複受精のもう1つの受精相手である中央細胞について、受精に伴うカルシウム濃度変化を調べました。すると、卵細胞と同様に、1回目のカルシウム濃度上昇は花粉管からの精細胞の放出と同時でしたが、2回目のカルシウム濃度上昇は中央細胞の受精のタイミングとは明確な一致が見られませんでした。中央細胞においては、卵細胞より複数の精細胞との受精が起こりやすいことが分かっており、この2回目のカルシウム濃度上昇が中央細胞の多精拒否に関わっている可能性も考えられます。
最後に、花粉管を誘引するのに重要な2つの助細胞についても、受精過程のカルシウム濃度変化を調べてみました。すると、花粉管が助細胞に接触した後、両方の助細胞でカルシウム濃度の振動が始まり、30分ほど持続した後、一方の助細胞で高いカルシウム濃度を示し、この助細胞は崩壊しました。崩壊時の助細胞のカルシウム濃度は最高値を示した後3分ほどで元の濃度に戻りましたが、驚いたことに、崩壊せずに残った助細胞が重複受精の完了後に再びカルシウム濃度の振動を示しました(図4)。花粉管の誘引停止に向け、残った助細胞で細胞死のプログラムが開始したことを示している可能性が考えられます。
重複受精に関わる4つの細胞の受精に伴うカルシウム濃度変化は、細胞によって異なっていていることが分かりました(図5)。重複受精を成功させるために、それぞれの細胞が役割を果たすためにカルシウム濃度変化によって細胞間のコミュニケーションをとりながら受精を進めていることが考えられます。
<今後の展開>
今回の研究成果により、受精に関わる4つの細胞それぞれの役割に応じた、特徴的なカルシウムシグナルの存在が明らかとなりました。重複受精における細胞間の情報伝達や、重複受精を制御する分子の解明に向けた重要な成果と言えます。卵細胞における受精のタイミング、つまり発生の開始点も明確に判断できるようになりました。今後は、こうしたカルシウムの挙動にどのような生理的意味があるのか調べることが焦点になります。細胞の中のカルシウム濃度を操作できる技術の開発が実現できれば、今回明らかになったカルシウム濃度変化が、重複受精においてどのような意味を持つのか理解することができます。それにより、2つの精細胞の受精相手が決まる仕組みが不明であるなど、いまだに多くの謎に包まれる植物の重複受精の仕組みが解明されることが期待されます。
<参考図>
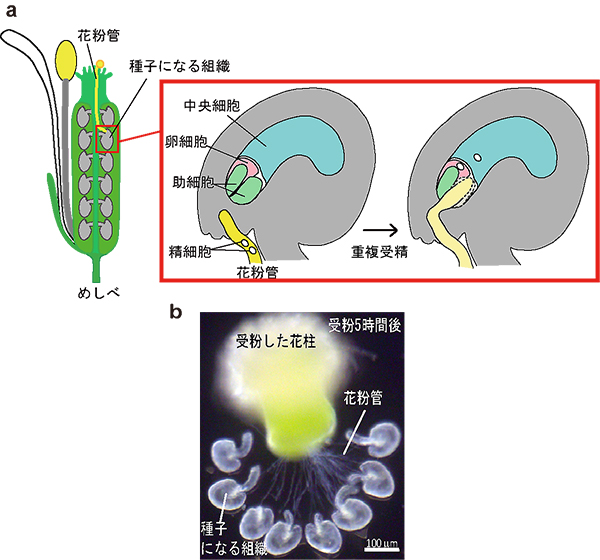
図1 シロイヌナズナの花および重複受精の模式図と体外重複受精系
aは、シロイヌナズナの花とその受精の様子を示しています。花粉は2つの精細胞を含み、花粉管を伸ばすことで精細胞を種子になる組織まで運びます。助細胞の誘引によって卵細胞の近くまで到達すると、花粉管は、精細胞を放出し、この時、2つある助細胞のうち一方が崩壊します。2つある精細胞のうち一方が卵細胞と受精して胚になり、もう一方は中央細胞と受精して胚乳になるこの被子植物の受精は、重複受精と呼ばれています。bは、卵細胞や中央細胞を種子になる組織から単離することなく、めしべから取り出して培地上で受精させる、シロイヌナズナ体外重複受精系を示しています。人工受粉した花柱を切り、培地上に置くと同時に、めしべから種子になる組織を取り出して培地に並べます。すると花柱から花粉管が伸び、受粉から約6時間後から受精を観察することができます。
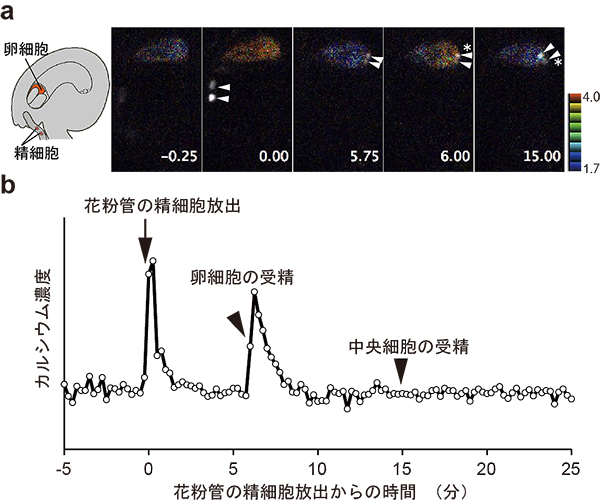
図2 卵細胞の受精に伴うカルシウム濃度変化
aはカルシウム濃度を擬似カラーで表示し、精細胞を白と矢尻で示してあります。*は受精した精細胞を示しています。bはaに示した例について、カルシウム濃度変化をグラフで表したものです。受精に伴って、卵細胞のカルシウム濃度の上昇が2回みられました。1回目は花粉管の精細胞放出のタイミングと一致していました。また、2回目のカルシウム濃度上昇は、卵細胞の受精のタイミングで起こりました。これに対して、中央細胞の受精の際には、カルシウム濃度変化はありませんでした。
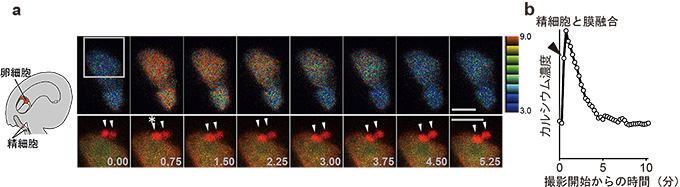
図3 卵細胞における2回目のカルシウム濃度上昇と精細胞の膜融合
aの上段では、カルシウムの濃度変化を擬似カラーで示し、下段では、上段の四角で囲った領域を拡大して、精細胞(矢尻)と卵細胞膜を赤で卵細胞の細胞質は黄色で示してあります。*は膜融合のタイミングを示しています。bは、aの例におけるカルシウム濃度変化をグラフに表したものです。卵細胞の2回目のカルシウム濃度上昇は、精細胞の膜融合のタイミングと一致していました。
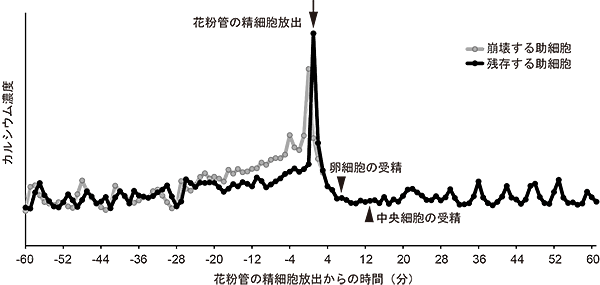
図4 助細胞における受精に伴うカルシウム濃度変化
2つの助細胞のカルシウム濃度変化をグラフで表したものです。灰色は花粉管が侵入して崩壊する助細胞を、黒色は花粉管の精細胞放出後も残存する精細胞について示しています。両方の助細胞で見られたカルシウム振動は30分ほど持続した後、崩壊する助細胞でより高いカルシウム濃度が維持されて、花粉管の精細胞放出でピークを迎えた後すぐに元のカルシウム濃度に戻りました。興味深いことに、卵細胞と中央細胞の受精後、残存している助細胞で再びカルシウム振動が見られました。
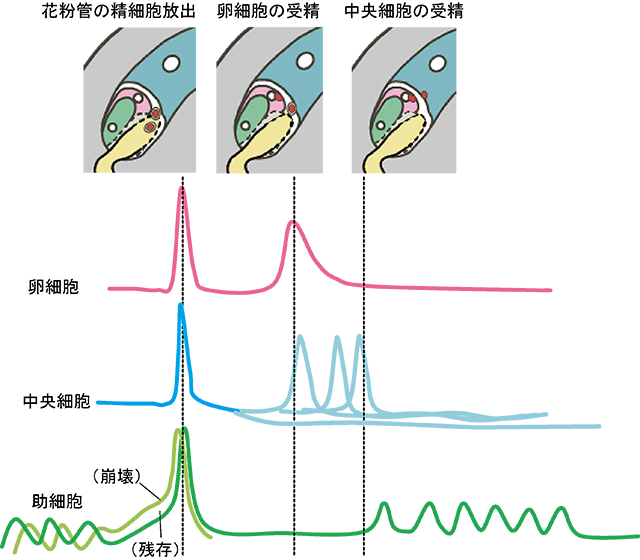
図5 重複受精に関わる4つの細胞の受精に伴うカルシウム濃度変化の模式図
花粉管が助細胞に誘引されて接触すると、2つの助細胞でカルシウム振動が起こりました。しばらく振動を続けると、次第に一方の助細胞でカルシウム濃度が高まり、花粉管による精細胞の放出が起こりました。この時、4つの全ての細胞でカルシウム濃度が最大値を示し、よりカルシウム濃度が高まっていた助細胞が一瞬で崩壊しました。そして約3分の間に全体でカルシウム濃度が下がると、次に卵細胞で単発のカルシウム濃度の上昇が見られました。この上昇は、卵細胞と精細胞の受精(細胞融合)に伴って起こることが明確に示されました。中央細胞では卵細胞のように受精に伴うカルシウム上昇は明確には観察されず、1回目のカルシウム上昇の後はさまざまなパターンのカルシウム上昇が観察されました。興味深いことに、卵細胞と中央細胞の受精後、残存している助細胞で再びカルシウム振動が見られました。
<用語解説>
- 注1) 膜融合
- 受精において、卵細胞(または中央細胞)と精細胞の細胞膜が融合すること。受精過程は、この細胞膜の融合の過程と、雌雄の核の融合の過程からなる。受精の瞬間という場合、一般に両者の細胞が融合する瞬間である膜融合の瞬間を指すことが多い。細胞壁を持つ植物においても、膜融合の起こる部位では細胞壁が存在しないと考えられており、植物においても受精は細胞膜の融合により進行する。
<論文名>
“Live imaging of calcium spikes during double fertilization in Arabidopsis”
(シロイヌナズナの重複受精におけるカルシウムスパイクのライブイメージング)
doi: 10.1038/ncomms5722
<お問い合わせ先>
<研究に関すること>
東山 哲也(ヒガシヤマ テツヤ)
名古屋大学 ERATO 東山ライブホロニクスプロジェクト 研究総括
名古屋大学 トランスフォーマティブ生命分子研究所 教授
〒464-8602 愛知県名古屋市千種区不老町
Tel:052-747-6404 Fax:052-789-2497
E-mail:
<JST事業に関すること>
中村 幹(ナカムラ ツヨシ)
科学技術振興機構 研究プロジェクト推進部
〒102-0076 東京都千代田区五番町7 K’s五番町
Tel:03-3512-3528 Fax:03-3222-2068
E-mail:
<報道担当>
科学技術振興機構 広報課
〒102-8666 東京都千代田区四番町5番地3
Tel:03-5214-8404 Fax:03-5214-8432
E-mail:
名古屋大学 総務部 広報渉外課
Tel:052-789-2016 Fax:052-788-6272
E-mail: