ポイント
- アブラナ科シロイヌナズナの突然変異体で、2本目の花粉管を高頻度に呼び寄せることを発見
- 卵細胞の隣にある2つ目の助細胞がカギ
- 農作物に1つでも多くの種子をつけたり、進化・生態学的な展開に期待
JST 課題達成型基礎研究の一環として、JST 戦略的創造研究推進事業 ERATO型研究「東山ライブホロニクスプロジェクト」(研究総括:東山 哲也 名古屋大学 大学院理学研究科 教授)の笠原 竜四郎 研究員らは、植物のめしべが受精に失敗した時に、積極的に受精を回復する仕組み(受精回復システム)が存在することを発見しました。
めしべに複数の花粉が受粉すると、複数の花粉管が一斉に伸長しますが、卵細胞には1本の花粉管だけが到達して受精することが知られています。動物のように多数の精子が卵に到達して競争の末に受精するケースとは異なり、植物では1本の花粉管、いわばたった1人の相手に受精を託すと考えられてきました。
笠原研究員らは、半数の花粉管が受精できない精細胞を持つシロイヌナズナの突然変異体を調べている時に、期待されるよりも多くの種子が作られることに注目しました。これまで、めしべの中の花粉管を全て観察することは困難でしたが、今回高度な解剖技術でこれを達成しました。その結果、これまではまれにしか起きないと考えられていた、卵細胞に2本の花粉管が到達する現象が、高頻度(約40%)で起こっていることを発見しました。さらに、1本目の花粉管が受精に失敗した場合に、卵細胞を含む組織が2本目の花粉管、いわば2人目の相手を呼び寄せ、受精を回復することを突き止めました。
多くの植物で、卵細胞の隣には助細胞という細胞が2つあり花粉管を誘引しますが、花粉管が到達すると助細胞の1つが崩壊し受精の場を作ります。これまで助細胞がなぜ2つあるのか、積極的な理由は見いだされていませんでした。今回、1本目の花粉管が受精に失敗した時に、2つ目の助細胞がバックアップ機能を果たし、受精を回復することも明らかにしました。
オオムギやエンドウなど、多くの植物で1つの卵細胞に花粉管が2本到達する例が確認されています。つまり、これまでまれなこととして注目されなかった現象が、植物生殖の重要な仕組みを意味していたと考えられます。今後この現象の分子メカニズムが解明されれば、植物の交配をコントロールできるようになり、例えば気象条件の厳しい地方で精細胞が原因で生じる農作物の不作に対して、1つでも多くの種子を形成させることができるようになると期待できます。
本研究成果は、2012年5月17日(米国東部時間)に米科学誌「Current Biology」のオンライン速報版で公開されます。
本成果は、以下の事業・研究プロジェクトによって得られました。
戦略的創造研究推進事業 ERATO型研究
| 研究プロジェクト |
「東山ライブホロニクスプロジェクト」 |
| 研究総括 |
東山 哲也(名古屋大学 大学院理学研究科 教授) |
| 研究期間 |
平成22年~平成27年 |
JSTはこのプロジェクトで、ライブセル解析による多細胞生物のホロニックコミュニケーションの全容解明に挑みます。多細胞生物の細胞は、種々のシグナル分子を介して、他の細胞と情報のやりとりをすることで自身の振る舞いを決定し、生物としてのバランスを取っています。こうした個と全体を調和する細胞間コミュニケーションを、ホロニックコミュニケーションと呼びます。そのメカニズムの解明および解析技術の確立は、広く生物学や医療の分野での貢献が期待できます。本研究では、そのための多くのライブセル解析技術の創出を目指します。
<研究の背景と経緯>
被子植物の精細胞は鞭毛を欠き、自ら泳ぐことはできません。このため精細胞は、花粉から伸びる「花粉管」により運ばれます。花粉管は卵細胞の隣に2つある助細胞により誘引され、卵細胞の近傍に到達します。花粉管が到達すると、花粉管の先端が破裂して精細胞が放出されます。それと同時に、一方の助細胞が崩壊して受精の場が作られます。この際、受精できない精細胞が放出されると、受精が成立しないため種子は形成されないと考えられてきました。しかし、これまで実際の様子は明らかにされませんでした。
笠原研究員らは、アブラナ科のシロイヌナズナを用いて受精に異常のある突然変異体を探索する中で、g21 という突然変異体を見いだしました。g21 変異体では半数の花粉で、受精できない精細胞が作られます。このことから、めしべの中には通常の半分(50%)の種子しかできないと予想されます。しかし、正確に種子の形成率(稔性)を調べたところ、g21 変異体の稔性が65~70%であることが明らかとなり、この観察が、本研究の発端となりました(図1)。
<研究の内容>
なぜg21 変異体で予想よりも多くの種子が作られるのか、詳細に調べることにしました。シロイヌナズナの1つのめしべには、およそ50個の卵細胞があります。その全てにおいて、花粉管が到達する様子を調べることができるように、解剖および観察技術を確立しました。その結果、g21 変異体の花粉を受粉しためしべでは、卵細胞には通常1本の花粉管しか向かわないとする定説に反し、約40%もの卵細胞で2本の花粉管が到達していました(図2)。2本目の花粉管が受精を回復しているのではないかと考え、次に最新のライブイメージング技術により、その瞬間をとらえることを試みました。その結果、1本目の花粉管が精細胞に異常を持ち受精に失敗した後、2本目の花粉管が正常な精細胞を放出し、受精が回復する様子をとらえることに成功しました(図3)。
次に、植物がどのように2本目の花粉管をコントロールしているのかを知るために、受精できない精細胞を持つ花粉管が特異的に染色されるようにし、その挙動を観察しました。その結果、1本目の花粉管が染色される場合に2本目の花粉管が到達し、1本目の花粉管が染色されない場合には2本目の花粉管は全く到達しないことが分かりました(図4)。これにより、めしべは受精が成立したか否かを感知し、積極的に2本目の花粉管を卵細胞まで誘引して受精の回復を試みるという仕組みが明らかとなり、この仕組みを「受精回復システム(Fertilization Recovery System)」と名付けました。
それでは、不運にも2本目の花粉管も受精に失敗した場合にはどうなるのでしょうか。興味深いことに、2本目が失敗すると、もはや種子を作れないことが分かりました。3本目の花粉管が到達して受精を回復することはなかったのです。このことは、卵細胞の隣にある助細胞という細胞の数と関係していると考えられます。助細胞は花粉管の誘引を担う重要な細胞ですが、花粉管が到達すると1つの助細胞が壊れて受精が行われます。2本目の花粉管が受精を試みる場合、2つ目の助細胞が1つ目と同様に花粉管を誘引したのちに壊れて受精の場を作ることが、先述のライブイメージングで確認されました。
本成果により、植物は、受精の失敗に備え、したたかに2つ目の助細胞をバックアップとして持っていると言えますが、やり直しのチャンスは一度限り、ということが分かりました。
今回の発見を以下にまとめます。
1.植物には、1本目の花粉管が受精に失敗した時に、2本目の花粉管を呼び寄せて受精する仕組みがある。
2.この仕組みにより、植物は受精を回復できる(受精回復システム)。
3.受精回復システムは、植物に助細胞が2つあるために成立する。
<今後の展開>
今後は、植物のこの受精回復システムがどのような分子メカニズムによって制御されているのかを解明していきます。植物の受精は、例えば高温、低温、多湿、乾燥といった厳しい気象条件に左右されることが知られていますが、この分子メカニズムが解明されれば、受精回復システムを人工的に促進することにより、農作物を含む植物に1つでも多くの種子を付けさせることが可能になるかもしれません。一方、自然界ではめしべに同種・異種のさまざまな花の花粉が運ばれることが知られています。異種の精細胞が1本目の花粉管で運ばれて受精に失敗した時に、2本目の花粉管を引き寄せることにより、同種の受精率の向上を図っている可能性が考えられます。このように、今回の発見により生態学分野でも新たな研究が進展することが期待されます。
さらには、多くの植物で助細胞がなぜ2つ存在するのかという疑問に対する答えの1つが示唆されたことにより、植物が雌の器官をいかにして発達させてきたかを知る進化学的な手掛かりにもなるのではないかと期待できます。
<参考図>
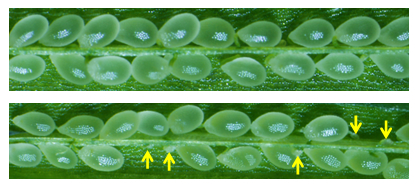
図1 g21 突然変異体における稔性
上の図は野生型のめしべ。全ての種子が形成されている。
下の図はg21 変異体(雄側の精細胞の変異による)のめしべ。矢印は形成不全の種子。理論的には50%の稔性になると予想されるが、正常種子の割合は形成不全の種子と比べて明らかに多く、本研究の発端となった。本研究で、g21 変異体だけでなく、既知の全ての突然変異体(精細胞の異常による受精不全)において、同様の稔性および受精回復システムが確認された。
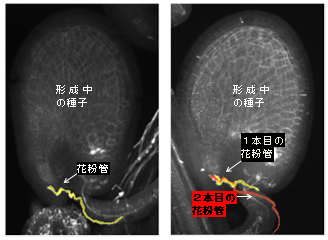
図2 二本の花粉管
左の図は野生型の胚珠(受精前の種子組織の名称)に花粉管が挿入されている様子。受精が完了し種子が形成されつつある。1つの胚珠には、卵細胞が1つと助細胞が2つ存在する。
右の図はg21 変異体の花粉を受粉した胚珠。2本の花粉管が観察できる。野生型と同様、種子が形成されつつある。花粉管が1本だけ到達して形成された種子と2本到達して形成された種子の割合は、めしべ内の全胚珠のうちのそれぞれ50%、18%であり、トータルすると種子の形成率68%となり、平均65%のg21 変異体の稔性とほぼ一致した。2本目の花粉管が、受精回復の要因であると考えられた。
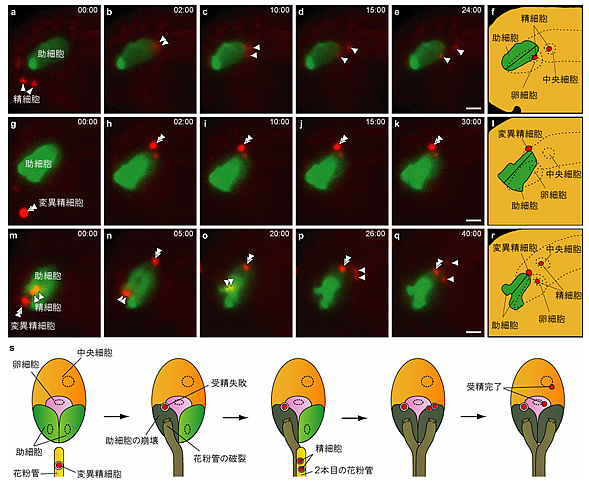
図3 受精回復システムのライブイメージング
a-f.野生型の受精の様子。助細胞を緑、精細胞を赤でラベルしてある。花粉管から勢いよく放出された精細胞のうち1つは卵細胞(受精すると胚になる細胞)に、もう1つは中央細胞(受精すると胚乳になる細胞)に受精している。
g-l.g21 変異体の精細胞が花粉管から放出された後の様子。変異精細胞は壊れた助細胞の端にとどまったまま受精することができない。
m-r.はじめにg21 変異体の精細胞が受精に失敗して、その後2本目の花粉管に由来する正常な精細胞が受精する様子。受精回復の瞬間がとらえられた。
s.受精回復システムの模式図。まず、変異精細胞を持つ花粉管が誘引されて助細胞に到達し、変異精細胞を放出するとともに、助細胞が崩壊する。しかしこの精細胞は受精に失敗し壊れた助細胞の端にとどまる。次に正常な精細胞を持つ花粉管が助細胞に到達し、助細胞を崩壊させながら精細胞を放出する。放出された精細胞はそれぞれ卵細胞と中央細胞に移動し、受精を完了させる。
この時、2本目の花粉管が仮に受精に失敗しても、花粉管を引き寄せる働きを持つ助細胞が2つとも壊れてなくなっているので、3本目の花粉管を同じ胚珠に誘引することはできない。
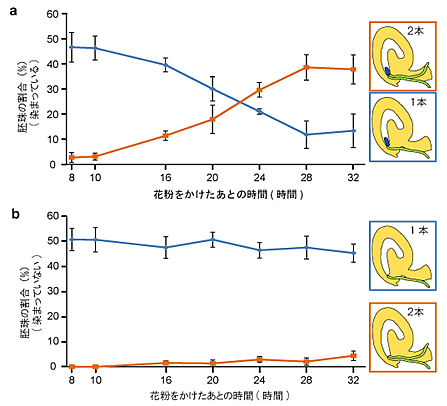
図4 受精に失敗した胚珠にのみ誘引される2本目の花粉管
a.花粉管が1本あるいは2本挿入されて青く染色されている胚珠(1本目の花粉管が受精を失敗した胚珠)の割合。青のグラフは花粉管を1本だけ受け入れた胚珠、またオレンジ色のグラフは2本受け入れた胚珠の割合を示している。花粉を受粉したあと10時間後までは胚珠は2本目を受け入れていないが、それ以降、28時間後までに40%の胚珠が2本目を受け入れていることが分かる。
b.花粉管が1本あるいは2本挿入されて青く染色されていない胚珠(1本目の花粉管が受精に成功した胚珠)の割合。受粉から32時間後に至るまで、2本の花粉管を受け入れた胚珠はほとんど見られない。
このことは、胚珠は受精に失敗した時のみ2本目の花粉管を誘引し、1本目の受精が成功した場合は2本目を誘引しないということを示している。また、1本目の花粉管が失敗してから2本目を受け入れるまでに10時間以上かかっていることから、胚珠が受精を回復するにはある程度の時間が必要であることも示唆している。
<論文名>
“Fertilization Recovery after Defective Sperm Cell Release in Arabidopsis”
(シロイヌナズナにおける受精能を欠く精細胞放出後の受精回復システム)
doi: 10.1016/j.cub.2012.03.069
<お問い合わせ先>
<研究に関すること>
笠原 竜四郎(カサハラ リュウシロウ)
名古屋大学 ERATO 東山ライブホロニクスプロジェクト 研究員
〒464-8602 名古屋市千種区不老町
Tel:052-747-6401 Fax:052-789-2497
E-mail:
<JSTの事業に関すること>
金子 博之(カネコ ヒロユキ)
科学技術振興機構 研究プロジェクト推進部
〒102-0075 東京都千代田区三番町五番町7 K’s五番町ビル
Tel:03-3512-3528 Fax:03-3222-2068
E-mail: