本研究成果のポイント
- B細胞が病原体成分に反応して抗体を産生するためにはCIN85が必須
- CIN85がB細胞抗原受容体からのシグナルを転写因子NF-κBへ伝達
- アレルギーや自己免疫疾患、リンパ腫の病態解明・治療への手がかり
独立行政法人 理化学研究所(理研、野依 良治 理事長)と国立大学法人 大阪大学(鷲田 清一 総長)は、免疫細胞の一種であるB細胞注1)が、肺炎球菌などの病原体成分に反応して抗体を産生するためには、抗原のシグナル伝達を担うアダプター分子注2)「CIN85」が必須であることを解明しました。これは、理研 免疫・アレルギー科学総合研究センター(谷口 克 センター長) 分化制御研究グループの黒崎 知博 グループディレクター(兼 大阪大学 免疫学フロンティア研究センター 分化制御研究室 特任教授)と米谷 耕平 研究員らを中心とする共同研究グループの成果です。
B細胞は、生体を脅かす細菌やウイルスなどの抗原に遭遇すると、抗体注3)を産生して抗原を攻撃し、排除します。そのため、B細胞が適切に活性化できないと免疫不全に陥ります。その反対に、B細胞が過剰に活性化するとアレルギーや自己免疫疾患、リンパ腫を引き起こす可能性があります。しかし、B細胞が活性化する仕組みはいまだに不明な点が多く、この仕組みを明らかにすることが急務とされていました。
研究グループは、B細胞内に存在し、細胞外からのシグナルの伝達を担うアダプター分子CIN85に注目し、このタンパク質がNF-κB注4)という転写因子注5)の活性化に必須であり、CIN85が存在しないと、病原体成分の一種であるⅡ型T細胞非依存的抗原(TI-Ⅱ抗原:ティーアイツー抗原)注6)に対して抗体を産生することができなくなることを発見しました。CIN85によるNF-κBの活性化の機構は、これまで知られていなかった全く新しいメカニズムです。
本研究は、JST 戦略的創造研究推進事業 チーム型研究(CREST)「アレルギー疾患・自己免疫疾患などの発症機構と治療技術」研究領域(菅村 和夫 研究総括)における研究課題「液性免疫制御による新しい治療法の開発」(研究代表者:黒崎 知博)の一環として行われたもので、米国科学雑誌「The Journal of Experimental Medicine」(2011年7月4日号)に掲載されるのに先立ち、オンライン版(米国東部時間:2011年6月27日付、日本時間:2011年6月27日)に掲載されます。
<背景>
私たちの体は、細菌やウイルスなどの異物(抗原)が侵入すると、体を防御するためにさまざまな反応を示します。B細胞による抗体の産生はその1つです。B細胞は、細胞の表面にあるB細胞抗原受容体(BCR)注7)で抗原を認識すると、細胞内に増殖、分化、活性化を引き起こすシグナルを伝達します。シグナルは多様な経路で伝わり、細胞の運命を決定する転写因子を活性化します。BCRからのシグナル伝達が欠如すると、B細胞は活性化できず抗体を作れなくなるため、免疫不全に陥ります。逆に、過剰な活性化はアレルギーや自己免疫疾患、リンパ腫といった疾患を引き起こす原因となります。そのため、BCRからのシグナル伝達経路を解明することは、これらの疾患の原因を突き止めたり、治療法を開発したりする上でも非常に重要です。
1998年に研究グループは、BLNKと呼ばれるアダプター分子が、BCRからのシグナルを伝達するために重要な役割を果たすことを突き止めていました。しかし、このBLNKが、その後どんな分子の修飾を受けてB細胞を活性化させるかについては不明のままでした。そこで、BLNKと結合するタンパク質を探索し、B細胞のシグナル伝達のメカニズム解明に挑みました。
<研究手法と成果>
研究グループは酵母を用いた分子生物学の手法により、BLNKにCIN85と呼ばれる別のアダプター分子が結合することを発見しました。そこで、このCIN85がB細胞のシグナル伝達において重要な機能を発揮しているのではないかと考え、B細胞だけでCIN85を欠損したノックアウトマウスを作製し、このCIN85欠損マウスと野生型マウスを比較しました。
まず、野生型マウスとCIN85欠損マウスにTI-Ⅱ抗原を投与し、産生する抗体の量を測定しました。TI-Ⅱ抗原は、肺炎球菌の細胞壁の一番外側の被膜(莢膜(きょうまく))に存在する、細菌成分に代表される抗原です。測定の結果、野生型では抗原を認識し、抗体を産生しましたが、CIN85欠損マウスでは抗体を産生しませんでした(図1)。このことは、CIN85が抗体産生に必須であることを示しています。
次に、細胞外からのどのような刺激がCIN85に作用するのかを突き止めるために、野生型マウスとCIN85欠損マウスのB細胞を精製し、B細胞を活性化させるさまざまな活性化因子を加えて試験管内で一緒に培養しました。その結果、野生型マウスのB細胞では、加えた全ての活性化因子に反応したのに対し、CIN85欠損マウスのB細胞では、BCRからの刺激に対して反応を示しませんでした(図2)。このことは、CIN85欠損マウスではBCRからのシグナルを伝達できず、免疫反応を引き起こすことができなかったことを示しています。
細胞内にはBCRからのシグナルを多方面へ伝えるさまざまなシグナル伝達経路が存在していますが、CIN85がどのシグナル伝達経路で働いているかを明らかにするために、詳細な生化学的解析を行ないました。その結果、CIN85が欠損していると、転写因子の1つであるNF-κBが活性化しないことが分かりました(図3)。
さらに、CIN85欠損マウスで認めた異常が、本当にNF-κBを活性化しないことが原因であると確認するために、CIN85欠損マウスにNF-κBを強制的に活性化させることのできる活性化型IKKβという分子を導入しました。その結果、活性化型IKKβを導入したCIN85欠損B細胞は、低下していたBCRに対する応答性を取り戻し、また、TI-Ⅱ抗原に対する抗体産生能も回復しました(図4)。
これらの実験結果から、CIN85欠損マウスが示した免疫応答の低下は、BCRからのシグナル伝達をCIN85が担えなくなり、NF-κBを活性化させられなくなったためであると証明できました。つまり、B細胞がTI-Ⅱ抗原に反応し抗体を作り出すためには、CIN85がBCRからのシグナルを転写因子NF-κBに伝達する必要があることが分かりました(図5)。
<今後の期待>
今回の結果から、CIN85が正しく機能しないと、肺炎球菌などの細菌に対する免疫応答を引き起こせなくなることが判明しました。このことは、免疫不全の原因がCIN85の機能不全にあるという可能性を示しています。逆に、もしCIN85が過剰に働いてしまうと、B細胞の異常な活性化を引き起こし、アレルギーや自己免疫疾患、リンパ腫などの疾患を引き起こす可能性も考えられます。特にNF-κBシグナル伝達経路では、その関連遺伝子の異常がリンパ腫を引き起こすことが知られており、創薬の面からも非常に注目されます。今回、CIN85によるBCRからNF-κBへのシグナル伝達経路を明らかにしたことにより、CIN85は免疫疾患の創薬開発における新たな標的分子となることが期待されます。
<参考図>
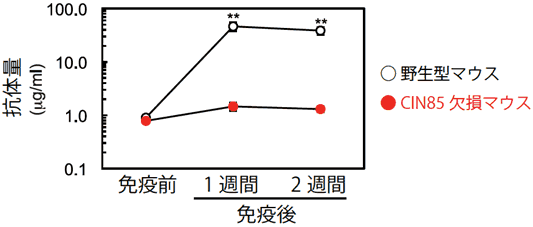
図1 CIN85欠損マウスはTI-Ⅱ抗原に対する抗体産生が低下している
抗体産生能を測定するために、野生型マウスあるいはCIN85欠損マウスにTI-Ⅱ抗原を投与した。免疫1週間後、2週間後に血清を採取し、抗原に対する抗体量を測定した。野生型マウスは免疫により抗体の産生が認められたが、CIN85欠損マウスでは認められなかった。この結果から、CIN85はTI-Ⅱ抗原に反応する上で必須であることが分かった。
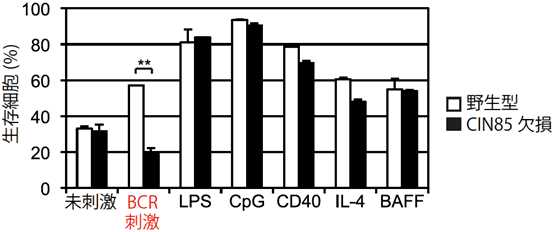
図2 CIN85欠損マウスのB細胞はBCRからの刺激に反応できない
細胞外からのどのような刺激に対してCIN85が働くのかを突き止めるために、B細胞をさまざまな活性化因子と共に試験管内で培養し、48時間後に生き残っている細胞の割合を測定した。野生型マウスのB細胞では、未刺激と比較して、全ての活性化因子で生存細胞数の割合が増加しているのに対し、CIN85欠損マウスのB細胞では、BCR刺激に対する生存細胞数が減少していた。このことから、CIN85はBCRからのシグナルを伝達するのに必須であることが分かった。
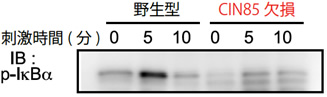
図3 CIN85欠損マウスのB細胞はNF-κBの活性化が低下している
CIN85欠損マウスが伝達するシグナル経路を同定するために、B細胞にBCRから刺激を加え、細胞内のタンパク質の活性化を測定した。野生型は刺激5分後に、NF-κBの活性化を示すp-IκBαリン酸化(黒いバンド)を認めたが、CIN85欠損では認められなかった。このことは、CIN85がNF-κBを活性化させるために必須な役割を果たしていることを示している。
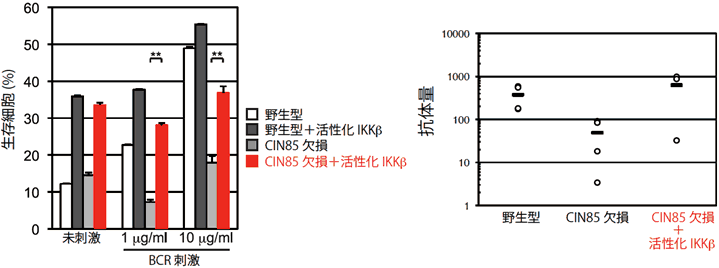
図4 CIN85欠損マウスで低下した免疫応答は活性化型IKKβで回復する
CIN85欠損マウスの表現型が、NF-κB経路のシグナル伝達の障害によるものであることを確認するために、直接NF-κBを活性化させることのできる活性化型IKKβをCIN85欠損マウスに導入して観察した。CIN85欠損マウスで認めた生存細胞数と抗体量の減少は、活性化型IKKβにより野生型と同程度にまで回復したことから、CIN85がBCRからのシグナル伝達をNF-κBに伝えていることを示している。
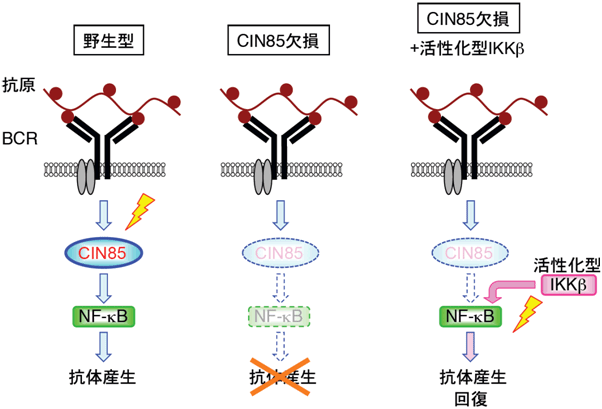
図5 CIN85はBCRからのシグナルをNF-κBに伝達する
- 左: 野生型では抗原によるBCRからのシグナルがCIN85を通じてNF-κBに伝達され抗体産生を引き起こす。
- 中: CIN85欠損ではBCRからのシグナルがNF-κBに伝達されず抗体産生が起きない。
- 右: CIN85欠損に活性化型IKKβを導入すると、CIN85が存在しなくてもNF-κB活性化され、抗体産生が回復する。
CIN85の働きが活性化型IKKβで代償できたことから、CIN85がNF-κBを活性化していることが分かった。
<補足説明>
- 注1) B細胞
- 免疫細胞の一種で、表面にあるB細胞抗原受容体(BCR、注7参照)と呼ばれるタンパク質で病原体などの抗原を認識し、抗体を産生する。
- 注2) アダプター分子
- 細胞表面の受容体と細胞内のシグナルとを橋渡しする役割を持つ分子のこと。アダプター分子自体は酵素活性を持たないが、種々のタンパク質結合構造を持ち、この構造にシグナル伝達分子が結合することで、細胞内シグナルネットワークに情報が伝わる。
- 注3) 抗体
- B細胞が分泌するタンパク質で、抗原に結合し生体防御に働く。抗体が結合した抗原は破壊され、体から排除される。
- 注4) NF-κB
- nuclear factor kappa Bの略で、核内転写因子の一種。活性化されると核内に移行し、染色体上の特異的なDNAモチーフに結合し、さまざまな遺伝子の転写を開始する。NF-κBは、免疫及び炎症反応、あるいは腫瘍形成の制御に重要な役割を果たす。
- 注5) 転写因子
- 遺伝子の働きをオンにしたりオフにしたりする機能を持つタンパク質。DNAの転写を制御する領域に結合し、遺伝子発現のタイミングや量を調節する。
- 注6) Ⅱ型T細胞非依存的抗原(TI-Ⅱ抗原)
- 多くの抗原に対しては、T細胞が寄与して抗体産生が誘導されるが、TI-Ⅱ抗原に対しては、T細胞の寄与がなくてもB細胞は活性化することができる。細菌の莢膜(きょうまく)に存在する多糖に代表されるように、繰り返し配列を持つことが特徴。
- 注7) B細胞抗原受容体(BCR)
- B細胞が細胞表面にもつ受容体。1つ1つのB細胞のBCRはそれぞれ異なった抗原特異性を持ち、無数ともいえる抗原に反応できる。
<お問い合わせ先>
<研究に関すること>
黒崎 知博(クロサキ トモヒロ)
独立行政法人 理化学研究所 免疫・アレルギー科学総合研究センター 分化制御研究グループ グループディレクター
国立大学法人 大阪大学免疫学フロンティア研究センター 分化制御研究室 特任教授
Tel:045-503-7085 Fax:045-503-7018
独立行政法人 理化学研究所 横浜研究推進部 企画課
Tel:045-503-9117 Fax:045-503-9113
<JSTの事業に関すること>
石井 哲也(イシイ テツヤ)
独立行政法人 科学技術振興機構 イノベーション推進本部 研究領域総合運営部
Tel:03-3512-3524 Fax:03-3222-2064
E-mail:
<報道担当>
独立行政法人 理化学研究所 広報室 報道担当
Tel:048-467-9272 Fax:048-462-4715
独立行政法人 科学技術振興機構 広報ポータル部
Tel:03-5214-8404 Fax:03-5214-8432
E-mail: