<本研究成果のポイント>
○Homerタンパク質が神経活動依存的にリン酸化されることを発見
○リン酸化が、Homer複合体のダイナミックな調節の契機
○長期記憶・学習を支えるシナプス後部の構造変化の包括的理解へ
独立行政法人 理化学研究所(野依 良治 理事長)と独立行政法人 科学技術振興機構(JST:北澤 宏一 理事長)は、シナプス後部に豊富に存在するタンパク質「Homer」のリン酸化が神経活動に伴うシナプス後部構造の再構築調節機構の一端を担うことを解明しました。理研脳科学総合研究センター(田中 啓治 センター長代行)発生神経生物研究チームの御子柴 克彦 チームリーダー、水谷 顕洋 研究員らの研究グループによって行われ、本研究成果の一部は、御子柴チームリーダーが研究代表者を務める「JST戦略的創造研究推進事業 発展研究 カルシウム振動プロジェクト」によって得られました。
私たちの大脳は、100億個以上の神経細胞から成り、それらが極めて複雑なネットワークを形成して、信号のやりとり(神経伝達)を行っています。この神経伝達は、神経細胞間の接点であるシナプスという特別な場所で行われていますが、信号の受け手側であるシナプス後部には、この信号を感知・解読し、それに対して反応するために、極めて多彩なタンパク質が配備されています。私たちの脳が記憶・学習するとき、反復する信号、あるいは異なる複数の信号が同時に1つの神経細胞に入ることで、ある特定のシナプスにおける神経伝達効率が強化された状態が維持されます。これを生み出している1つのメカニズムが、シナプス後部のタンパク質群の質的・量的変化だと考えられていますが、その詳細な分子メカニズムはほとんどわかっていません。
Homer(ホーマー)タンパク質は、シナプス後部に豊富に存在する
アダプター分子注1)で、シナプス後部で種々のシグナル分子間の複合体形成を担っています。研究グループは、小脳の
Purkinje(プルキンエ)細胞注2)のシナプス後部に特異的に存在するHomer3に着目し、Homer3が神経活動に依存してリン酸化され、それにより、Homer3と代謝性グルタミン酸受容体との複合体が消失することを発見しました。さらに、このHomer3リン酸化が、Purkinje細胞シナプス後部でのカルシウムシグナリングを調節する可能性を示しました。この発見は、Homerタンパク質が、そのリン酸化というタンパク質修飾反応を利用して、シナプス後部の分子複合体構造を神経活動に対して極めて柔軟に調節する分子であることを世界で初めて示したものです。これは、記憶・学習を支えるシナプス後部の構造変化を包括的に理解する上で、大変重要な知見となります。
本研究成果は、米国の科学雑誌『
Journal of Neuroscience』(5月14日号)に掲載されました。
<1.背景>
私たちの大脳は、100億個以上もの神経細胞から成り立っています。また、1個の神経細胞は、約1万個もの神経細胞とシナプスという部分で手をつなぐことが可能です。神経細胞は、このように極めて複雑なネットワークを形成し、信号(神経活動)のやりとりを行っています。例えば、外界の状況変化は、感覚神経によって感知された信号として脳に入ります。脳内に入った信号は、一群の神経細胞の興奮を契機として、複雑なネットワーク上を伝搬しますが、その過程で私たちは外界を感知し、それに対する反応・行動を決定します。
記憶・学習は、反復する信号、あるいは異なる複数の信号が同時に1つの神経細胞に入ることで、ある特定のシナプスにおける神経伝達だけが強化されて起こると考えられています。神経細胞間の神経伝達の本体は、シナプス前部からグルタミン酸を放出し、それによるシナプス後部のグルタミン酸受容体の活性化であることが知られています。実際に、記憶・学習に伴ったこれらグルタミン酸放出とグルタミン酸受容体の質的・量的変化については、これまでにも数多くの報告がなされています。しかし、記憶・学習が長期にわたって保持されるためには、このグルタミン酸放出とグルタミン酸受容体の変化だけでは不十分で、グルタミン酸受容体を取り巻くシナプス後部の多彩なタンパク質の分子的構造変化が、特定のシナプスにおいてのみ長期にわたって維持される必要があります。この長期間の記憶・学習を支えるシナプス後部で起きている詳細な分子メカニズムは、依然として不明のままとなっていました。
Homerは、シナプス後部に豊富に存在するタンパク質で、Homer1、Homer2、Homer3の3種類の仲間が知られています。共通する特徴は、自身が4量体を形成することで、Homer結合タンパク質同士を会合させる、いわばアダプターとして働くことです。代謝性グルタミン酸受容体、IP3受容体、SHANK注3)、drebrin注3)などは、いずれもシナプス後部に局在する、Homer結合タンパク質として知られています。これら結合タンパク質は、Homerを介してシナプス後部で機能的複合体を形成し、シナプス後部のカルシウムシグナリングや細胞骨格アクチンの制御に関与しています。上述のように、長期記憶・学習に際しては、シナプス後部の分子的構造変化が必須であるにも関わらず、Homerタンパク質を介した複合体に関して、神経活動に反応して起こるダイナミックな変化は全く知られていませんでした。研究グループは、Homer3が小脳Purkinje細胞に特異的に発現していること(図1左)に着目し、Homer3を介する複合体の神経活動依存的なダイナミックな変化を捉えることに挑みました。その結果、Homerタンパク質のリン酸化によるシナプス後部分子構造調節、という全く新しい機能を発見することになりました。
<2.研究手法と成果>
(1)リン酸化Homer3の発見
生化学的手法を用いて、マウスの小脳をシナプス後部画分と可溶性画分とに分離調製したところ、Homer3の一部が可溶性画分にも存在することが判明しました。さらに、可溶性画分中のHomer3を2次元電気泳動にて分離解析したところ、1分子あたり少なくとも3カ所がリン酸化していました。
この結果は、本来シナプス後部にあるべきHomer3が、リン酸化によって大きく性質を変え、可溶性になったことを意味します。すなわち、このリン酸化が、長期記憶・学習を確立する上で極めて重要な分子構造変化を調節していると考えられました
(図1右)。
(2)Purkinje細胞内のCaMKIIがHomer3のリン酸化酵素
そこで、Homer3のリン酸化を行っているリン酸化酵素の同定を試みたところ、以下の4つの結果を得ました。
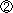
リン酸化セリン残基特異的抗体で認識すると、Purkinje細胞内に3カ所のセリン残基がリン酸化されたHomer3が存在する。
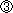
初代培養Purkinje細胞を脱分極刺激すると、Homer3がリン酸化される。一方、CaMKII阻害剤「KN-93」を加えるとリン酸化が阻害される
(図2)。
これらの結果から、CaMKIIがPurkinje細胞内でHomer3をリン酸化する酵素であると結論づけました。
(3)リン酸化によるHomer3と結合タンパク質との親和性低下
Homer3がリン酸化されるとHomer3は可溶性になり、リン酸化がHomer3の結合タンパク質との親和性を低下させる可能性があると考えました。そこで、リン酸化していないHomer3とリン酸化したHomer3とを試験管内リン酸化実験で準備し、それぞれの結合タンパク質に対する結合動態を、表面プラズモン共鳴原理を利用した解析装置で調べました。その結果、リン酸化Homer3は、非リン酸化Homer3に比べて約10倍も結合親和性が低下していることがわかりました
(図3)。実際、初代培養Purkinje細胞において、脱分極刺激によるCaMKIIのHomer3リン酸化によって、Homer3と代謝性グルタミン酸受容体との複合体が消失することを発見しました。
(4)Homer3を介する複合体のリン酸化によるダイナミックな調節
小脳Purkinje細胞のシナプス後部に存在するHomer3を介する代謝性グルタミン酸受容体-Homer3-IP3受容体複合体に対する、Homer3のリン酸化による機能的影響を検討しました。具体的には、特殊な増殖細胞であるHeLa細胞を用いた一過性発現系を用いて、この複合体のアウトプットであるカルシウムシグナルについて解析しました。その結果、Homer3のリン酸化によってこの複合体が極めてダイナミックに調節されていることを見いだしました。
(5)まとめ
Homer3は、Purkinje 細胞のシナプス後部において、神経活動に伴って活性化されたCaMKIIによってリン酸化されると、その結合タンパク質との結合親和性が低下します。これは、Homer3を介した複合体の「柔軟化」をもたらし、それにより、その複合体の神経活動依存的機能変化を引き起こします。Homer3のリン酸化が、神経活動に伴ったシナプス後部の分子構造の柔軟化を担っている、と結論することができます。
<3.今後の期待>
神経活動に反応するリン酸化反応は、一般的に極めて早い反応で、実際、Homer3のリン酸化も刺激後、2分後から検出できます(図2左)。こうした変化が、長期間の記憶・学習にとってどういう意味を持つのかが問題になります。研究グループは、この神経活動早期に、神経伝達の入力を受けたシナプス後部においてのみ起きるHomer3のリン酸化と、それによるシナプス後部の分子構造の柔軟化を、長期記憶・学習確立の契機となる出来事だと考えています(図4)。現在、この考えを実証するために、Homer3のリン酸化部位を破壊したノックインマウスの作製に挑んでいます。
<報道担当・問い合わせ先>
(問い合わせ先)
独立行政法人 理化学研究所 脳科学総合研究センター 発生神経生物研究チーム
チームリーダ― 御子柴 克彦(みこしば かつひこ)
Tel:048-467-9745 Fax:048-467-9744
脳科学研究推進部 企画課
大伴 康志(おおばん やすし)
Tel:048-467-9596 Fax:048-462-4914
(JSTの事業に関すること)
独立行政法人 科学技術振興機構 戦略的創造事業本部 研究プロジェクト推進部
小林 正(こばやし ただし)
Tel:03-3512-3528 Fax:03-3222-2068
(報道担当)
独立行政法人 理化学研究所 広報室 報道担当
Tel:048-467-9272 Fax:048-462-4715
独立行政法人 科学技術振興機構 広報・ポータル部 広報課
Tel:03-5214-8404 Fax:03-5214-8432
<補足説明>
注1)アダプター分子
種々の機能タンパク質が複合体を形成し機能する時、これら機能タンパク質同士を会合させるのに必要な仲介的分子を言う。機能タンパク質同士を会合させるのに必要なモデュールを複数個有するか、Homerタンパク質のようにそれ自身が多量体を形成することによって、複合体形成に関与する。
注2)Purkinje(プルキンエ)細胞
小脳皮質に存在する唯一の出力細胞で、小脳が関与する記憶・学習において、中心的な働きをする神経細胞である。
注3)SHANK、Drebrin
いずれも、シナプス後部を形成する骨格タンパク質である。
注4)CaMKII(カルシウムカルモデュリン依存性キナーゼII)
神経細胞、特に、シナプス後部に豊富に発現するタンパク質リン酸化酵素。細胞内のカルシウムイオン上昇に伴って活性化することで、種々のタンパク質をリン酸化する。記憶・学習に深く関わっていることが知られている。
注5)P/Q タイプ電位依存性カルシウムチャンネル
Purkinje細胞のシナプス後部に豊富に発現するカルシウムチャンネルで、神経活動に伴って起こるPurkinje細胞シナプス後部での細胞外からのカルシウムイオン流入をつかさどる。
<参考図>
図1 Homer3のマウス小脳Purkinje細胞における分布とそのリン酸化
(左)小脳Purkinje細胞におけるHomer3の発現と分布を示す。Homer3はPurkinje細胞の細胞体から樹状突起にかけて広汎に分布している。特に、樹状突起上のシナプス部位であるスパインに強く分布している。
(右)小脳Purkinje細胞におけるリン酸化Homer3の分布を示す。リン酸化Homer3が、細胞体と樹状突起の基幹部分により強く分布しているのがわかる。
図2 初代培養Purkinje細胞でのCaMKIIによるHomer3のリン酸化
(上)初代培養Purkinje細胞をCaMKIIの阻害剤(KN-93)存在下と非存在下(vehicle)で、脱分極刺激した際のHomer3のリン酸化の様子を、リン酸化Homer3に結合するpSer抗体による染色像で示す。脱分極後、KN-93非存在下では2分後に顕著にリン酸化が起きているのがわかる(左半)。このリン酸化が、KN-93存在下では阻害されることから、CaMKIIがHomer3のリン酸化酵素であると結論できる。
(下)上段と同じ試料のHomer3抗体による染色像である。
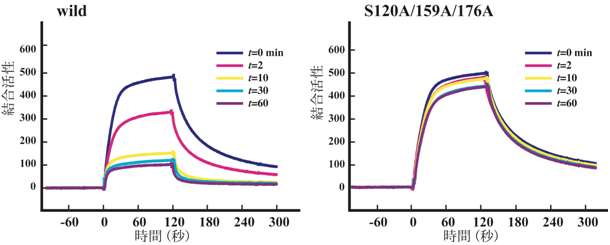
図3 リン酸化によるHomer3結合タンパク質に対する親和性の低下
(左)Homer3 野生体を示した時間(0分、2分、10分、30分、60分)、CaMKIIで試験管内リン酸化し、それぞれの時間におけるHomer結合タンパク質との結合動態を解析した。時間が経過し、リン酸化反応が進むにつれて、結合親和性が減少する。
(右)左と同様の実験を3つのリン酸化セリン残基を全てアラニンに置換したHomer3変異体(S120A、159A、176A)で行うと、時間が経過してもリン酸化反応が進まないため、結合親和性の減少が見られない。
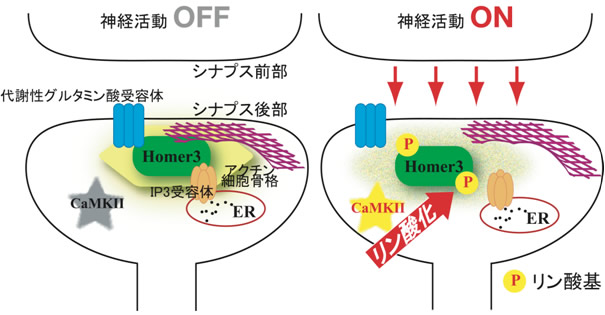
図4 シナプス後部におけるHomer3リン酸化の機能
シナプス後部では、Homer3を介してHomer結合タンパク質同士が複合体を形成している。神経活動が入ると(ON)、シナプス後部のCaMKIIが活性化し、Homer3をリン酸化する。これによってHomer3と結合タンパク質との結合が弱くなり、この複合体が解離する。こうしてHomer3のリン酸化によってシナプス後部の分子構造が「柔軟」になることが、長期記憶・学習確立のための重要な出来事と推測している。