| ■ | 体の非対称 |
| 「対称な形態から、いかにして非対称が生じるのか?」すなわち「体の軸はいかにして形成されるのか?」哺乳類では、胚発生の途中で初めて非対称性が生じる。一体、どのような機構で非対称性が生じるのだろうか? |
| ■ | 体の非対称性はどのようにして生じるのだろうか? |
| 濱田研究代表者らのグループは、これまで体の左右非対称性ができる仕組みを研究してきた。この左右非対称に関しては、我々は数々の成果を発表しており、例えば、平成8年には、哺乳類で非対称に発現する初めての遺伝子として、レフティ(Lefty)(※1)と呼ばれる遺伝子の役割などを明らかにし報告した。
今回は前後、すなわち頭尾方向の形成の仕組みの解明に挑戦した。哺乳類の場合、対称な位置にあった遠位臓側内胚葉(DVE distal visceral endoderm)(※2)と呼ばれる特殊な細胞が片方へ移動し、その結果、移動した側に頭が形成されるという現象が既に知られていた。しかしながら、何故、DVEが片側(将来の頭側)へ移動するのか、細胞が移動する方向はどのように決められているのか、といった細胞が移動するメカニズムについては不明のままであった。今回の研究では、これらの細胞移動のメカニズムの問題を解明しようとした。 |
| ■ | 頭尾の方向の決定:図1参照 |
| マウスでは、受精後5.5~6.5日の間に頭尾軸が確立する。これにはいくつかのステップがある。まず、受精後5.5日には胚の最も遠位側(胎盤から遠い位置)に、DVEと呼ばれる特殊な細胞集団が生じる。この細胞DVEは、遠近軸以外の方向性に関して対称に位置している。次に、DVEが胚の片側(将来の頭側)へ移動し始める。片側へ移動したDVEは、頭側を誘導するシグナル因子レフティ1(Lefty1)やサール(Cerl)を分泌し、DVEが移動した側を頭、反対側を尾とする。しかしながら、DVEが移動する仕組みについては全く知られていなかった。
今回、マウス胚を用いて以下の様な実験を行い、DVEがなぜ頭側へ移動するかという謎を解く重要な知見が得た。
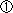 |
レフティ1(Lefty1)の発現を調べたところ、DVEが移動を始める前にすでに片側に偏って発現していた。レフティ1(Lefty1)と同様に、ノーダル(Nodal)の抑制因子として働くサール(Cerl)も同様に非対称に発現していた。 |
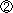 |
DVEが移動を始める前のマウス胚へレフティ1(Lefty1)やサール(Cerl)の機能を導入すると、DVEは導入された側へ移動した。 逆に、ノーダル(Nodal )を導入すると、導入した場所とは逆の方向へ移動した。 |
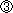 |
ノーダル(Nodal )はDVEを含む臓側内胚葉(VE: visceral endoderm)の細胞分裂を上昇させ、レフティ1(Lefty1)やサール(Cerl)は逆に抑制した。 |
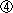 |
レフティ1(Lefty1)やサール(Cerl)を欠損するマウスでは、DVEの移動が大幅に遅れた。(それでも移動をしたのは、代償的に発現したレフティ2(Lefty2)のためである。) |
|
| ■ | 左右と頭尾の決定機構には、同じ分子が関与 |
| 以上の結果から、レフティ1(Lefty1)やサール(Cerl)の非対称な発現が、細胞分裂を非対称性に抑制することにより、DVEの移動する方向、ひいては体の頭尾の方向を決めていることがわかった。左右の決定で重要な役割を持つレフティ(Lefty)やノーダル(Nodal)が、もう一つの体の軸である頭尾軸をも決めていることになる。左右と頭尾の決定機構は、細胞レベルでは全く違うメカニズムでありながら、同じ分子が関与するというのは興味深い。
また、発生途中の細胞が移動することは、体が出来上がる過程において、様々な場所で認められているが、細胞分裂速度の違いが移動方向を決めるというメカニズムはまだ他に例がない。 |
| ■ | 社会的意義 |
| 我々の体は外見的に左右対称であるが、内臓の臓器の多くは3次元的に非対称である。例えば、肝臓は方向性を持った非常に複雑な形状を示している。このような複雑な臓器を作るためには多様な種類の細胞が必要であるが、それに加えて、細胞集団内に方向性(非対称性)が存在することが必須である。今回、我々は左右に加えて、前後の非対称性の機構に大きな知見を得た。本研究を進めていくことで、生物の前後・左右を決定する遺伝子プログラムの全体像に迫りたい。本研究が発展していくことで、体が出来上がるための基本的な仕組みが解明されると期待される。
更に、本研究で得られた知識は、将来の医療として注目される再生医療、とくに複雑な形態と機能を持つ臓器を再生するためには重要になるであろう。また、ヒトの先天性奇形の原因究明やその早期治療法確立につながることも期待される。 |