ポイント
- 植物の雄(花粉管)が雌(卵細胞)を探すための誘引物質を感知する受容体を発見した。
- 複数の受容体が協調して雌のシグナルを感知し、花粉管を伸長させることを解明した。
- 受精の仕組みの解明の他、種子生産の効率化や異種間受精の技術開発が期待される。
JST 戦略的創造研究推進事業において、ERATO東山ライブホロニクスプロジェクトの東山 哲也 研究総括(名古屋大学 WPI トランスフォーマティブ生命分子研究所 教授)と光技術グループの武内 秀憲 博士研究員(現所属:オーストリア グレゴール・メンデル研究所)らは、被子植物の雄である花粉管が、雌である卵細胞を見つけるために用いる受容体キナーゼ注1)を発見しました。
花粉管は、自身を伸長させることで雌しべの中を進み、卵細胞を含む雌しべの奥深くの最終目的地まで精細胞を運ぶという、受精するための重要な役目を担っています。東山教授らは、花粉管を誘導するために雌しべが分泌する誘引物質「ルアー」を発見し、ルアーが植物の種類によって異なり、同種の花粉管のみを呼び寄せることを明らかにしてきました。しかし、花粉管がルアーを感知する仕組みは分かっていませんでした。
本研究では、シロイヌナズナの花粉管の先端部分で、ルアーを感知する受容体を発見しました。この受容体は、他のよく似た複数の受容体と協調して働くことで、雌しべ組織からのシグナルを正確に感知していることも分かりました。これらの受容体キナーゼが雌しべのさまざまなシグナルを受け取ることで、ルアーを感知できる距離まで花粉管を伸長させ、花粉管は雌しべの中を目的地まで迷わずに進んでいることを明らかにしました。
本研究成果は、植物が同種間で受精するための仕組みの全容解明に大きな前進をもたらしました。発見した受容体の研究が進めば、受精効率を操作して種子生産の効率を高める技術や、異種間受精を容易にする手法の開発につながると期待されます。
本研究成果は、2016年3月10日(英国時間)に英国科学誌「Nature」に掲載されます。
本成果は、以下の事業・研究プロジェクトによって得られました。
戦略的創造研究推進事業 総括実施型研究(ERATO)
| 研究プロジェクト |
「東山ライブホロニクスプロジェクト」 |
| 研究総括 |
東山 哲也(名古屋大学 世界トップレベル研究拠点プログラム(WPI) トランスフォーマティブ生命分子研究所 教授) |
| 研究期間 |
平成22年10月~平成28年3月 |
多細胞生物の細胞が種々のシグナル分子を介して、他の細胞と情報のやり取りをすることで自身の振る舞いを決定し、生物としてのバランスを取るための仕組み「ホロニックコミュニケーション」の全容解明に向けた、多くのライブセル解析技術の創出を目指します。
<研究の背景と経緯>
私たちが普段食べている米や大豆は植物の種子です。野菜も種子から成長することで収穫できるようになります。植物が種子を生産するためには、雄と雌が出会い、受精に成功する必要があります。被子植物の雄は、花粉とその内部に含まれる精細胞からなっています。花粉は管状の単一細胞からなる花粉管へと発達し、その先端部分を伸長させることで雌しべの中を進んでいきます(図1A)。花粉管は最終的に卵細胞を含む雌しべの奥深くの目的地まで到達し、精細胞を卵細胞へと受け渡すことで受精が達成されます。このように、花粉管が迷うことなく卵細胞を見つけていることが、私たちの食料生産を支えているといえるかもしれません。雄と雌が迷わずに出会うという神秘的で重要な現象ですが、その仕組みは多くの謎に包まれています。
2009年に東山教授らは、卵細胞の隣にある助細胞が花粉管を誘導するために分泌する誘引物質「ルアー」をトレニアで発見しました。2012年には、シロイヌナズナでもルアーを発見し、ルアーは植物の種類によって少しずつ異なり、他種の花粉管ではなく同種の花粉管を呼び寄せることで、同種間での正常な受精が保証されていることも明らかにしました(図1B)。ルアーこそが、雄を呼び寄せるために雌が分泌している鍵因子であることが分かりました。一方で、花粉管がどのようにルアーを感知しているのか、どのようにルアーを感知できる距離まで進んでいるのか、伸長や誘引に関わる花粉管側の仕組みはほとんど知られていなかったため、ルアーを認識する花粉管側の鍵因子を一から発見する必要がありました。
<研究の内容>
モデル植物であるアブラナ科のシロイヌナズナを用いて、ルアーを感知する花粉管側の鍵因子を探索しました。ルアーを感知する鍵因子候補として、花粉管の膜表面に局在する受容体キナーゼが考えられました。候補とした23個それぞれの受容体キナーゼの機能が損なわれた花粉管に対して、ルアーを用いたバイオアッセイ注2)を行いました。その結果、ルアーの感知に必要不可欠な受容体PRK6注3)を発見することに成功しました(図2)。
PRK6には他に、花粉と花粉管だけで働く、アミノ酸配列がよく似たファミリーの受容体注4)が複数存在します。そこで、他の複数のPRK受容体の機能を損なわせたところ、PRK受容体の組み合わせにより花粉管がルアーに反応しにくくなったり、花粉管の伸長が悪くなったりしました。これは、雌しべからのシグナルがPRK受容体に作用することで花粉管の伸長が促進されるという過去の他の研究グループの報告とも一致する結果です。このことから、ルアーの感知だけでなく、花粉管がルアーを感知できる距離まで効率的に伸長するためにも、PRK6とその他のPRK受容体が協調して働いていることが分かりました。
次に、PRK6が花粉管の細胞内にどのようなシグナルを伝えることで、ルアーに反応しているのかを調べました。花粉管が真っすぐ伸長している時、細胞膜上のPRK6は左右均等に局在しています。花粉管先端で蛍光標識したPRK6をイメージング解析したところ、ルアーを与えると、ルアーの側の細胞膜上にPRK6が集まり、集まった方向に花粉管が伸長方向を変えていく様子が観察されました(図3)。このことから、PRK6が花粉管の伸長に重要な因子群をルアーの方向に集めることで、ルアーに向かって伸長していくという仕組みが考えられました。
花粉管の誘引は同種間だけで働く仕組みです。シロイヌナズナと同じアブラナ科のナズナ(ぺんぺん草)注5)に対してシロイヌナズナのルアーを用いたバイオアッセイを行ったところ、通常のナズナはシロイヌナズナのルアーに反応しませんでした。そこで、ナズナの花粉管にシロイヌナズナのPRK6の遺伝子を導入して同様に実験しました。興味深いことに、遺伝子の導入によりシロイヌナズナのPRK6を発現するようになったナズナの花粉管はシロイヌナズナのルアーに応答できるようになりました(図4)。PRK6受容体が同種のルアーを認識する鍵因子であることが分かりました。
本研究の成果を以下にまとめました。
- 1) 花粉管が誘引物質ルアーを感知するのに必要不可欠な受容体を世界で初めて発見しました。
- 2) 複数のPRK受容体が協調して働くことで、雌しべ組織からのさまざまなシグナルを受け取り、ルアーを感知できる距離まで花粉管を伸長させていることを明らかにしました。
- 3) 同種に対して働くシロイヌナズナのルアーをPRK6が中心となって感知していることを示しました。
<今後の展開>
植物の雄しべを雌しべに受粉させると種子ができることは紀元前から知られている身近な現象で、農業にも重要な仕組みです。また、花粉管が雌しべ組織に誘導されることは100年以上も昔に発見されており、花粉管を伸長させること自体は容易にできるため、現在では中高校生の実験課題にもなっています。世界的には、誘引物質ルアーが発見されたことで、その受容と応答の仕組みの解明が待たれていました。本研究で花粉管の伸長とルアーの感知を担う花粉管側の鍵因子を発見したことで、花粉管の効率的な伸長とルアーに反応するための仕組みの一端が明らかとなりました。PRKファミリー受容体に着目した今後の研究で、花粉管の効率的な伸長や同種のルアーの感知による受精の仕組みの全容解明が期待されます。
また、本研究では、PRK6受容体の遺伝子導入により異種の花粉管を誘引できることが分かりました。この知見を生かして研究を進めれば、異種間受精を可能とする方法の開発につながるかもしれません。さらに、受容体は一般的にさまざまな薬剤の標的となり得ます。PRK受容体を標的とした薬剤を探索することで、受精効率を操作して種子生産の効率を高めたり異種間受精を容易にしたりするような薬剤の開発が期待できます。
<参考図>
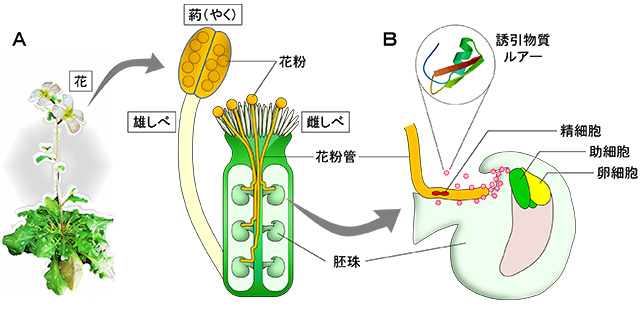
図1 シロイヌナズナの花粉管の伸長と誘引物質ルアーによる誘導
- A) シロイヌナズナの花および雄しべと雌しべの模式図。雄しべ先端の葯(やく)の中で作られた花粉は、受粉すると花粉管を伸長させて雌しべの中を進んでいく。最終的に花粉管は卵細胞を含む、受精により種子へと発達する組織(胚珠)に到達する。
- B) 卵細胞の隣にある助細胞が誘引物質ルアーを分泌し、花粉管を誘導する。助細胞に到達した花粉管は精細胞を放出し、卵細胞へと受け渡すことで受精が行われる。シロイヌナズナのルアーは70個程のアミノ酸からなるたんぱく質で、シロイヌナズナの花粉管に対して効率的に働く。図のような構造をしていると予想されている。

図2 ルアーを感知できないprk6変異体の花粉管とPRK6受容体の模式図
- A) 野生型の花粉管は、培地上に置いたルアーを含むゼラチンで作ったビーズ(星印)に向かって伸長の方向を変えた(左パネル)。一方、PRK6受容体を作ることができない変異体(prk6変異体)の花粉管はルアーに全く反応することはなかった(右パネル)。PRK6がルアーの感知に必要不可欠であることが分かった。矢尻は花粉管の先端を示す。
- B) PRK6受容体たんぱく質の模式図。細胞外にロイシンリッチリピートドメイン注6)を、細胞内にキナーゼドメインをそれぞれ持つ、膜貫通型のたんぱく質である。
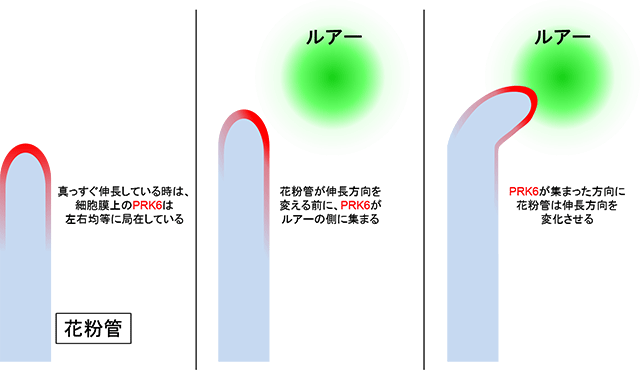
図3 ルアーに応答したPRK6の局在変化
PRK6は花粉管の先端の細胞膜上に主に局在している。ルアーに応答して花粉管が伸長方向を変える(形を変化させる)前に、PRK6は伸長していく方向に集まることが分かった。PRK6が花粉管の伸長に重要な因子群をルアーの方向に集めることで、伸長方向を決定していると考えられる。

図4 ナズナへのシロイヌナズナPRK6の導入実験
実験に用いたナズナ(ルベラナズナ)の花の写真と花粉管の誘引実験。シロイヌナズナのルアーを含むビーズ(星印)に、通常のナズナは誘引されないが、シロイヌナズナのPRK6を導入したナズナの花粉管は誘引された。矢尻は花粉管の先端を示す。
<用語解説>
- 注1) 受容体キナーゼ
- 動物の受容体キナーゼと似た構造を持つたんぱく質のファミリーで、受容体様キナーゼともいう。数多くの遺伝子ファミリーが見つかっており、細胞膜を1回貫通し、細胞外の領域を持つタイプのものだけで、シロイヌナズナは450遺伝子以上持っている。細胞内の領域には、たんぱく質のリン酸化に関わり、一般的には細胞内にシグナルを伝えるために重要とされるキナーゼドメインを持つ。
- 注2) ルアーを用いたバイオアッセイ
- 培地上に伸長させた生きた花粉管にルアーを作用させて、反応するかどうかを調べる実験。図2と図4は、実際の様子である。
- 注3) PRK6
- Pollen-specific Receptor-like Kinase 6を表す略称(和訳:花粉特異的受容体様キナーゼ)。PRK6は、シロイヌナズナでは8つ存在するPRKと名付けられたたんぱく質の一群の1つである。過去の研究からそのうちのいくつかが花粉管の伸長に関与することが報告されてきたが、実際の雌しべ内での重要性は全く分かっていなかった。
- 注4) よく似たファミリーの受容体
- 受容体キナーゼは、たんぱく質のアミノ酸配列を比較することで、いくつかのグループに分類される。さらに8つのPRKのように、似たもの同士を一群の仲間としてより細かく分類できる。よく似た受容体たんぱく質の一群は、それぞれが同じ機能を持つ場合もあれば、少しずつ異なる機能を持つ場合もある。
- 注5) ナズナ(ぺんぺん草)
- 日本でも春に道端でよく見かけるナズナ(ぺんぺん草)。研究に用いたのは、その仲間でありゲノム情報が解読されているルベラナズナ(学名 Capsella rubella)である。モデル植物であるシロイヌナズナ(学名 Arabidopsis thaliana)に近縁な種であることから、近年、さまざまな比較解析に用いられている。
- 注6) ロイシンリッチリピートドメイン
- アミノ酸のロイシンを多く含むたんぱく質の構成単位の1つ。さまざまなたんぱく質で見られ、たんぱく質間の相互作用にもしばしば関与する。
<論文タイトル>
“Tip-localized receptors control pollen tube growth and LURE sensing in Arabidopsis ”
(シロイヌナズナの花粉管先端に局在する受容体群が花粉管の伸長とルアーの感知を制御する)
doi :10.1038/nature17413
<お問い合わせ先>
<研究に関すること>
東山 哲也(ヒガシヤマ テツヤ)
ERATO 東山ライブホロニクスプロジェクト 研究総括
名古屋大学 WPI トランスフォーマティブ生命分子研究所 教授
〒464-8602 愛知県名古屋市千種区不老町
Tel:052-747-6404 Fax:052-747-6405
E-mail:
<JST事業に関すること>
大山 健志(オオヤマ タケシ)
科学技術振興機構 研究プロジェクト推進部
〒102-0076 東京都千代田区五番町7 K’s五番町
Tel:03-3512-3528 Fax:03-3222-2068
E-mail:
<報道担当>
科学技術振興機構 広報課
〒102-8666 東京都千代田区四番町5番地3
Tel:03-5214-8404 Fax:03-5214-8432
E-mail:
名古屋大学 総務部 広報渉外課
〒464-8601 愛知県名古屋市千種区不老町
Tel:052-789-2699 Fax:052-788-6272
.
E-mail: