ポイント
- 植物の体内時計は組織を単位として半ば独立に生理応答を制御していることを示した。
- 植物の温度情報の処理における、表皮の体内時計の重要性を初めて示した。
- 植物組織の体内時計機能は、植物の精密な成長調節法開発のターゲットとして期待される。
JST 戦略的創造研究推進事業において、京都大学の 遠藤 求 准教授らは、植物の体内時計の働きを組織レベルで阻害する手法を用いて、各組織における体内時計の生理的意義を明らかにすることに成功しました。
本研究グループの先行研究から、動物のような体内時計注1)の機能分担が、植物にも組織レベルで存在することは示されていましたが、どの組織の体内時計がどのような生理応答に関わっているかは不明でした。
本研究グループは、体内時計への代表的な入力刺激である日の長さ(日長)と温度がそれぞれどの組織の体内時計によって処理されているか解析した結果、維管束(篩部)の体内時計は日長情報を処理し花芽形成(花成)を制御する一方で、表皮の体内時計は温度情報を処理し細胞伸長を制御していることを明らかにしました。このことは、植物の体内時計は各組織の体内時計が半ば自律的に制御を行う非集中型のネットワーク構造をとっていることを示しており、植物の体内時計の機能分担が、より明確になりました。
さらに、今回の結果は植物に対する温暖化の影響を評価する上で表皮を標的とすることの重要性を示しているばかりでなく、未だ発見されていない植物の温度受容体が表皮に存在する可能性を強く示唆するものです。より精度の高い1細胞レベルでの解析や特定の組織だけを標的とした成長制御により、植物の精密な成長調節法の開発につながると期待できます。
本研究成果は、2015年11月2日(英国時間)に英国科学誌「Nature Plants」のオンライン速報版で公開されます。
本成果は、以下の事業・研究領域・研究課題によって得られました。
戦略的創造研究推進事業 個人型研究(さきがけ)
| 研究領域 |
「細胞機能の構成的な理解と制御」
(研究総括:上田 泰己 東京大学 大学院医学系研究科 教授) |
| 研究課題名 |
構成的アプローチによる植物の生物時計の組織特異的な役割の解明 |
| 研究期間 |
平成23年12月~平成26年11月 |
<研究の背景と経緯>
これまで、植物の体内時計は個々の細胞が独自に時間を測っており、どの組織の体内時計も同じ機能を持っていると漠然と考えられていたため、植物個体全体を用いた解析が主流であり、植物組織ごとの体内時計機能や組織・細胞間での体内時間情報のやりとりについてはほとんど解析されていませんでした。遠藤准教授らは昨年、植物の花成制御に維管束の体内時計が重要であることを世界に先駆けて明らかにしましたが(Endo et al., Nature 2014)、維管束の体内時計が植物の生理応答全般に重要であるかどうか、また、温度など他の環境情報の処理にも関わっているのか、については未解明のままでした。
また、日長と温度は植物にとって特に重要な環境情報です。特にストレスではない温度(常温)に対する植物の応答を正確に予測することやそのシグナル伝達経路を明らかにすることは、農業上の課題であるだけでなく、地球温暖化が植物に及ぼす影響を評価する上で極めて重要です。しかし、日長を測る光受容体や受容組織が明らかにされているのに対して、温度受容体や受容組織は未だ明らかにされていないため、植物の温度シグナル伝達がどこでどのように起こっているかは不明のままでした。
温度や日長は体内時計の入力シグナルとして機能していることから、体内時計を介した植物の環境感覚の仕組みを明らかにすることができれば、これらの問題の解決に大きく前進すると予想されました。
<研究の内容>
体内時計はほぼ全ての細胞・組織で見られるため、変異体の解析や全身で過剰発現した植物個体の解析からは、観察された表現型がどの組織の体内時計の機能に由来しているのかを推定するのは困難でした。遠藤准教授らはまず、各組織の体内時計がどのように環境情報を処理しているか明らかにするため、モデル植物であるシロイヌナズナ注2)を用いて、組織ごとに体内時計機能を阻害した系統を作出しました。これにより、観察される表現型と組織の機能の関係がはっきりとし、初めて定量的に各組織の体内時計の機能を解析することが可能になりました(図1)。これらの系統を用いて、花芽形成を誘導する長日条件および、誘導しない短日条件での、花芽形成速度を測定したところ、維管束の特に篩部伴細胞注3)で時計機能を阻害した系統のみが長日条件下で遅咲きを示すことがわかりました(図2)。さらに、この篩部伴細胞の時計は花成ホルモン(フロリゲン)注4)をコードしている遺伝子の転写を制御するものの、上流の遺伝子も含め、日周変動には大きな差異が見られなかったことから、篩部伴細胞の時計は、日長依存的な花芽形勢誘導の制御を、極めて限られた標的の制御を通じて行っていることが考えられました。
次に、他の生理応答でもこうした維管束・篩部伴細胞の時計が同様に重要であるかを調べるために、代表的な体内時計制御の生理応答である胚軸伸長制御について調べました。維管束・篩部伴細胞の時計遺伝子の過剰発現系統では全く表現型が見られない一方で、表皮の時計機能を阻害した系統では顕著な胚軸注5)伸長が観察されました(図3)。さらに、こうした胚軸伸長制御は、約17℃〜27℃のこの植物にとっての常温の範囲でのみ観察されたことから、表皮の時計は温度情報の処理に関わっていることがわかりました(図4)。その一方で、こうした表皮の時計による胚軸伸長制御は日長の影響を受けませんでした。
また、表皮の時計は胚軸以外にも子葉や葉柄の伸展・伸長に関わっていたことから、表皮の時計は細胞伸長制御全般を司っていることが示されました。こうしたことから、維管束の時計が日長情報を処理し花芽形成を制御しているのに対して、表皮の時計は温度情報を処理し、細胞伸長制御に関わっていることがわかりました。
こうした結果は、維管束・篩部伴細胞の時計と表皮の時計はそれぞれ異なる環境情報を処理し、異なる生理応答の制御に関わっていることを示しています。しかし、これらの時計は完全に独立というわけではなく、ゆるく相互作用していることが示唆されています。こうした体内時計のネットワーク構造は非集中型として知られているものであり(図5)、多様な環境でたくましく生きる植物の生存性略をよく反映しているものでした。こうしたネットワーク構造が明らかになったことで、植物を組織・細胞レベルのネットワークとして理解する必要性が示されました。
<今後の展開>
組織や細胞ごとに体内時計は役割が大きく異なることが本研究から明らかになったため、花芽形成や細胞伸長など体内時計によって制御されている生理応答の解析も、組織レベルそして1細胞レベルで行っていく必要があることがわかりました。
さらに、組織間での時間情報のやりとりが想定されますので、組織間シグナル伝達についても本研究で開発した手法を用いることで、こうした組織レベルでの解析が大きく進むことが期待されます。
また、維管束の時計機能を操作すると花芽形成を、表皮の時計機能を操作すると細胞伸長を独立して制御できたことから、体内時計を標的とした遺伝子組み換え植物の作出など、新たな植物の成長調節法開発が期待されます。
こうしたことに加え、温度情報の処理に表皮が重要であることが初めて明らかになったことから、今回の結果は、未知の温度受容体の探索の重要な指標になるだけでなく温度変化に対する植物の応答の正確な計測・予測など、進みつつある温暖化が植物に与える影響を評価するうえで重要な知見になると期待されます。
<参考図>
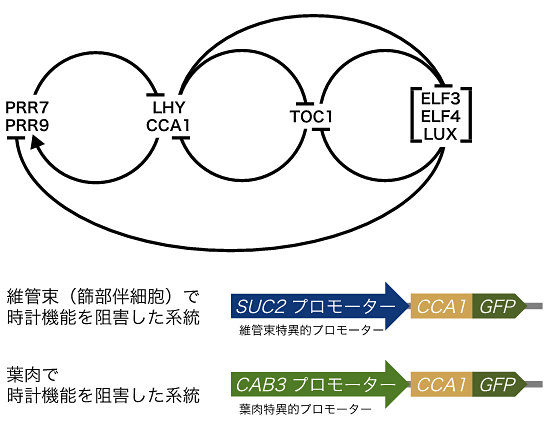
図1 組織特異的に体内時計機能を阻害する方法
- 上: 各アルファベットはシロイヌナズナの時計遺伝子名。ELF3、ELF4、LUXは複合体を形成し機能する。矢印は転写促進を「T字」は転写抑制を示す。植物の体内時計は転写・翻訳を伴うフィードバック・ループで構成されているため、時計遺伝子の過剰発現はリズムを失わせる。
- 下: このことを利用し、組織特異的な発現を持つことが知られているプロモーター注6)の下流に時計遺伝子(CCA1-GFP)をつなぎ、野生型に組織特異的に時計遺伝子を過剰発現させることで、組織特異的に時計機能を阻害した系統を作出した。
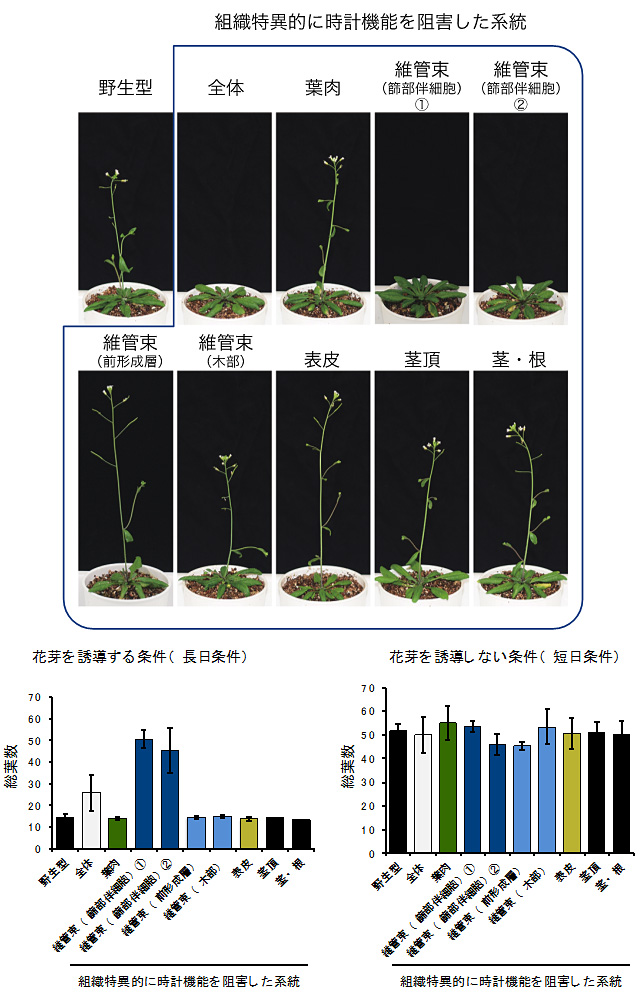
図2 維管束・篩部伴細胞の時計は日長依存的に花芽形成を制御する
- 上: 花芽を誘導する日長条件(長日条件)における花芽形成時期。野生型で花が咲く時期になっても、維管束・篩部伴細胞を含む組織で時計機能を阻害した系統では花が咲かない。ここでは、花芽形成速度を総葉数で計測しており、葉数が多いほど遅咲きであることを示している。
- 下: 花芽を誘導しない日長条件(短日条件)では維管束・篩部伴細胞の時計の効果が見られないことから、維管束・篩部伴細胞の時計は日長依存的に機能し、花芽形成を制御している。

図3 表皮の時計は細胞伸長を制御する
表皮を含む組織で時計機能を阻害した系統でのみ、22℃で胚軸(茎)が伸びている。花芽形成に表現型がみられた維管束・篩部伴細胞の時計は細胞伸長に影響しない。
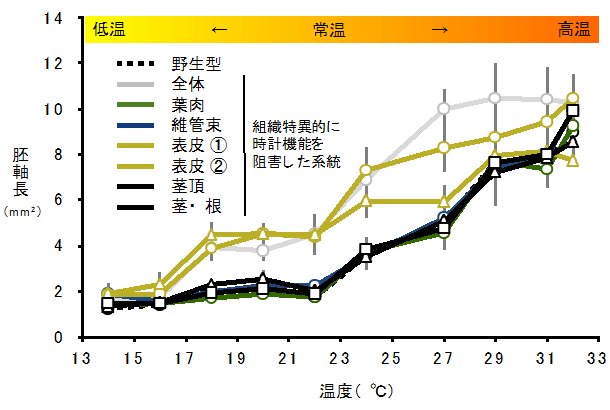
図4 表皮の時計による胚軸伸長制御は温度依存的である
表皮の時計を阻害した系統(黄色)および全体の時計を阻害した系統(灰色)では、17℃〜27℃の常温の範囲で野生型やその他の系統より胚軸が長い。このことから、表皮の時計は温度依存的に細胞伸長を制御している。
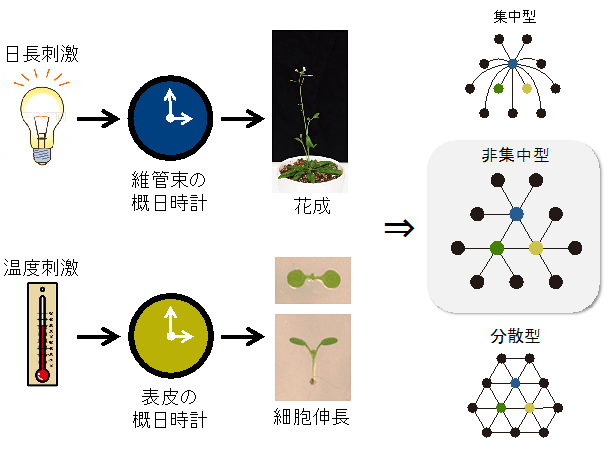
図5 今回明らかになった、植物の体内時計の組織特異的な機能分担とそのネットワーク構造
維管束の概日時計は日長情報を処理することで花芽形成を制御する一方で、表皮の概日時計は温度情報を処理して細胞伸長を制御している。独立した環境情報を独立した組織の時計が処理し、それぞれ異なる生理応答を制御していることから、植物の体内時計のネットワーク構造は集中型ではなく非集中型であることが明らかとなった。分散型のネットワークは階層構造がないネットワーク構造であり、特定の組織の時計機能を阻害しても表現型が現れないと考えられることから、今回は該当しない。色付きの丸は集中型もしくは非集中型でハブとなりうる組織を、黒丸はそれによって制御される組織・細胞を表している。
<用語解説>
- 注1) 体内時計
- 約24時間周期のリズムを生み出すための仕組み。地球の自転や公転に伴う明暗周期や季節変化に対応するための仕組みとして獲得されたと考えられている。
- 注2) シロイヌナズナ
- 代表的なモデル植物。アブラナ科。多くの時計遺伝子がすでに同定されており、体内時計の仕組みの解明が最も進んでいる。
- 注3) 篩部伴細胞
- 維管束は、栄養を輸送する篩管、水を輸送する道管、分裂組織を含む形成層からなっている。篩管はさらに、篩管篩部伴細胞と篩管細胞に大きく分けられ、篩部伴細胞は篩管細胞の機能を制御している。
- 注4) 花成ホルモン(フロリゲン)
- 植物の花芽形成(花成)を誘導する植物ホルモン。FTタンパク質がその実態である。
- 注5) 胚軸
- 芽生えの茎であり、子葉と根をつないでいる部分。
- 注6) プロモーター
- DNAにおける転写制御を担う領域であり、シロイヌナズナには組織ごとに発現することが知られている組織特異的プロモーターが数多く報告されている。
<論文タイトル>
“Decentralized circadian clocks process thermal and photoperiodic cues in specific tissues”
(非集中型の体内時計は温度刺激および日長刺激を特定の組織で処理している)
doi :10.1038/nplants.2015.163
<お問い合わせ先>
<研究に関すること>
遠藤 求(エンドウ モトム)
京都大学 大学院生命科学研究科 分子代謝制御学 准教授
Tel:075-753-6142 Fax:075-753-6470
E-mail:
<JST事業に関すること>
鈴木 ソフィア沙織(スズキ ソフィアサオリ)、川口 哲(カワグチ テツ)、山岸 裕司(ヤマギシ ユウジ)
科学技術振興機構 戦略研究推進部 ライフイノベーショングループ
〒102-0076 東京都千代田区五番町7 K’s五番町
Tel:03-3512-3525 Fax:03-3222-2064
E-mail:
<報道担当>
科学技術振興機構 広報課
〒102-8666 東京都千代田区四番町5番地3
Tel:03-5214-8404 Fax:03-5214-8432
E-mail: