ポイント
- ヒトiPS細胞注1)からヒト始原生殖細胞注2)を効率よく誘導する方法の開発に成功。
- ヒト始原生殖細胞の形成にBLIMP1 注3)が重要な役割を果たすことを証明。
- ヒトとマウスの生殖細胞形成機構の違いを解明。
- ヒト生殖細胞発生メカニズム解明の基盤を形成。
京都大学 大学院医学研究科の斎藤 通紀 教授(兼 京都大学 物質-細胞統合システム拠点(iCeMS=アイセムス) 主任研究者、京都大学 iPS細胞研究所 研究員)と同研究科の 佐々木 恒太郎 特定研究員、横林 しほり 特定助教らの研究グループは、ヒトiPS細胞からヒト始原生殖細胞様細胞を効率よく誘導する方法論の開発に成功しました。
これまで本研究グループは、マウスを用いて生殖細胞の発生機構を解明してきました。それらの知見に基づき、マウスES細胞注4)(mESC)やマウスiPS細胞から始原生殖細胞をサイトカイン注5)により誘導し、さらに精子、卵子を作製することに成功してきました。
今回の研究では、ヒトiPS細胞からヒト始原生殖細胞を効率よく誘導する方法の開発に成功しました。ヒトiPS細胞はプライム型の多能性注6)を有し、生殖細胞への誘導は難しいと考えられてきました。しかし、研究グループは、ヒトiPS細胞を特定のサイトカイン等で処理することにより、初期中胚葉様細胞(incipient mesoderm-like cells:iMeLCs)注7)に誘導し、さらにそれらをマウス始原生殖細胞の誘導と同様の方法で誘導することで、ヒトやカニクイザル注8)の始原生殖細胞とよく似た遺伝子発現を示すヒト始原生殖細胞様細胞(human primordial germ cell-like cells:hPGCLCs)が、効率よく誘導されることを証明しました。また、研究グループは、この方法を用いて、ヒト始原生殖細胞の形成にBLIMP1 が重要な役割を果たすこと、ヒトとマウスの生殖細胞形成機構に様々な違いが存在することを証明しました。この研究により、ヒト生殖細胞発生メカニズム解明の基盤が形成され、ヒト生殖細胞の発生機構の解明が大きく前進すると期待されます。
本研究成果は、JST 戦略的創造研究推進事業の一環として行われ、2015年7月16日正午(米国東部時間)に米国科学誌「Cell Stem Cell」のオンライン速報版で公開されます。
ヒトES細胞やヒトiPS細胞等からの生殖細胞の作成には、国の指針注9)に基づき、機関内倫理審査委員会による審査と、文部科学大臣への届け出が必要です。
本成果は、以下の事業・研究領域・研究課題によって得られました。
戦略的創造研究推進事業
| 研究プロジェクト |
「斎藤全能性エピゲノムプロジェクト」 |
| 研究総括 |
斎藤 通紀(京都大学 大学院医学研究科 教授) |
| 研究期間 |
平成23年度~平成28年度 |
<研究の背景と経緯>
生殖細胞とは、それらの融合により新しい個体を形成する精子や卵子、ならびにそれらを生み出す一連の細胞群です。本研究グループは、マウスを用いて、精子や卵子の起源となる始原生殖細胞(Primordial Germ Cells:PGCs)の発生機構を研究してきました。研究グループは、マウスES細胞やマウスiPS細胞などの多能性幹細胞から、サイトカインを用いて、エピブラスト様細胞(Epiblast-like cells:EpiLCs)注10)、さらには始原生殖細胞様細胞(PGC-like cells:PGCLCs)を誘導することに成功しました。この始原生殖細胞様細胞からは精子や卵子を分化誘導可能であり、さらにそれらを用いて子孫を得ることもできました。研究グループは、この培養技術を用いて生殖細胞の形成機構やエピゲノムリプログラミング注11)機構を解明してきました。
その一方で、マウスES細胞やiPS細胞はナイーブ型の多能性注6)を有するのに対し、ヒトES細胞やiPS細胞はプライム型の多能性を有しているため、生殖細胞への人為的な誘導は難しいと考えられてきました。
<研究の内容>
研究グループは、ゲノム編集技術注12)を用いて、2つの遺伝子BLIMP1 とTFAP2C 注13)が発現するとそれぞれ赤色、緑色の蛍光を発するヒトiPS細胞を樹立しました。BLIMP1 やTFAP2C は、ヒト始原生殖細胞で発現すると予想される転写制御因子注14)です。研究グループはそのiPS細胞を用いて始原生殖細胞を誘導する培養条件を検討しました。その結果、iPS細胞をサイトカイン等で処理することにより、まず、初期中胚葉様細胞(iMeLCs)に誘導し(図1)、さらにマウス始原生殖細胞の場合と同様の方法で処理するという、マウスとは異なる細胞状態を経由させることでBLIMP1 やTFAP2C を高く発現するヒト始原生殖細胞様細胞(hPGCLCs)が効率よく誘導されることがわかりました(図2、図3)。
こうして得られたヒト始原生殖細胞様細胞は、ヒトやカニクイザルの始原生殖細胞とよく似た遺伝子発現パターンを示し、初期のヒト始原生殖細胞に似た状態であることが示唆されました。ヒト始原生殖細胞様細胞は、マウスと同様、生殖細胞の発生過程で特徴的に見られるエピゲノムリプログラミングを起こしますが、マウスの始原生殖細胞とは遺伝子発現パターンが大きく異なることもわかりました。研究グループは、ヒト始原生殖細胞の形成に転写制御因子BLIMP1 が重要な役割を果たすこと、また、ヒトとマウスの生殖細胞形成機構に様々な違いが存在することを証明しました。さらに、あらかじめゲノム編集をiPS細胞に用いなくても、2種類の細胞表面マーカー注15)で生きた細胞を標識することで、ヒトiPS細胞から誘導したヒト始原生殖細胞様細胞を高い純度で単離できることも証明しました。これにより、原則的には、どのiPS細胞からもヒト始原生殖細胞様細胞を誘導・単離することができるようになりました。
<今後の展開>
この研究により、ヒト生殖細胞発生メカニズム解明の基盤が形成され、これまで非常に困難であったヒト生殖細胞の発生機構の解明が大きく前進すると期待されます。ヒト始原生殖細胞からヒト精子やヒト卵子の誘導が可能となれば、ヒトの遺伝情報継承機構の解明が進むのみならず、不妊症や遺伝病の発症機序解明に役立つと期待されます。
また、ヒトES細胞やヒトiPS細胞はプライム型の多能性を有し、その分化能は、ナイーブ型の多能性を有するマウスのES細胞やiPS細胞よりも低いと考えられてきました。本研究により、ヒトiPS細胞から生殖細胞が誘導されることが証明されたことから、ヒトiPS細胞も、基本的には、マウスES細胞/iPS細胞と同様の高い分化能を有することが示されたことになり、ヒトiPS細胞を用いた分化誘導研究一般の促進にもつながることが期待されます。
<参考図>
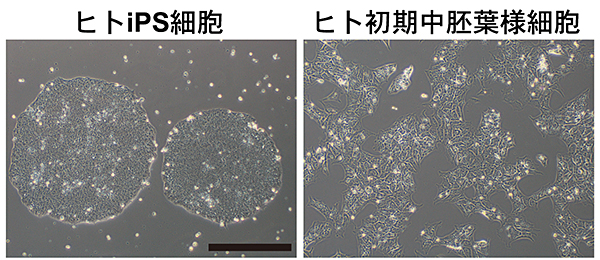
図1 ヒトiPS細胞とそれから誘導したヒト初期中胚葉様細胞
ヒトiPS細胞とそれから誘導したヒト初期中胚葉様細胞の位相差顕微鏡写真。
スケールバー:200μm
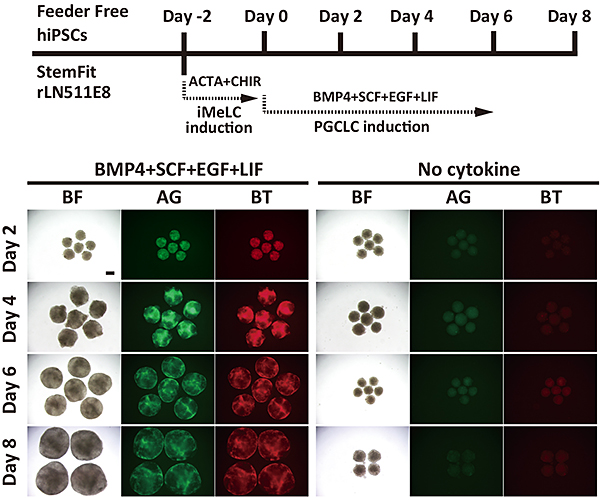
図2 ヒトiPS細胞からのヒト始原生殖細胞様細胞の誘導
ヒトiPS細胞をヒト初期中胚葉様細胞に誘導し(48時間の誘導)、それらを用いて浮遊細胞塊を形成させ、特定のサイトカインを作用させると、BLIMP1(赤色の蛍光)及びTFAP2C(緑色の蛍光)を発現するヒト始原生殖細胞様細胞が誘導される。
スケールバー:200μm
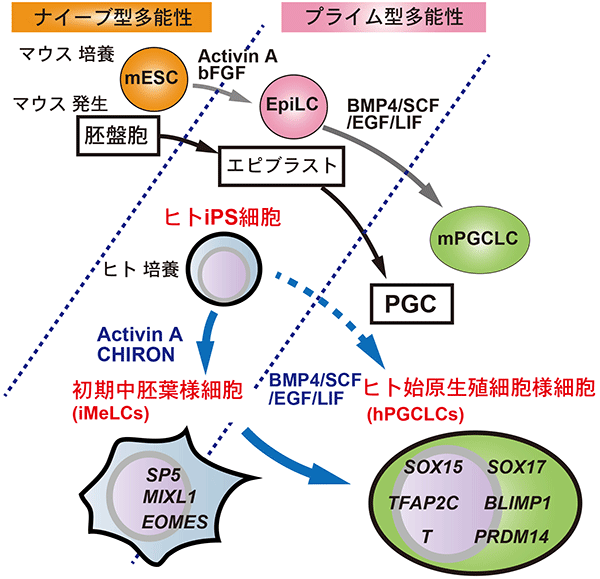
図3 ヒト始原生殖細胞様細胞の誘導モデル
これまではヒトiPS細胞から始原生殖細胞様細胞を直接誘導することは困難だった。今回、初期中胚葉様細胞を経ることでヒトの始原生殖細胞様細胞を誘導できることを示した。
<用語解説>
- 注1) iPS細胞
- 人工多能性幹細胞(induced Pluripotent Stem Cells)のこと。皮膚などの体細胞に特定の因子(Oct4,Sox2,cMyc,Klf4 など)を導入することにより作製される。ES細胞(注4参照)のように、無限に増殖し、生殖細胞を含むほぼ全ての組織に分化することができる。
- 注2) 始原生殖細胞
- 卵子もしくは精子の起源となる細胞。マウスの場合は胚齢6.5日前後に、胚体外胚葉(エピブラスト)(注10参照)内に誘導される。胚齢12日目前後に体細胞の性によって、卵子もしくは精子へと分化する。ヒト始原生殖細胞の発生機構はほとんど不明である。
- 注3) BLIMP1
- BLIMP1 はマウス始原生殖細胞の形成に必須な転写制御因子(注14参照)のひとつ。始原生殖細胞における体細胞化の抑制、多能性の再獲得、エピゲノムリプログラミング(注11参照)全てに重要な役割を果たす。ヒト始原生殖細胞の発生におけるBLIMP1 の機能は不明であった。
- 注4) ES細胞
- 受精卵から作製される胚性幹細胞(Embryonic Stem Cells)のこと。マウスの場合は受精後3-4日目の胚盤胞の内部細胞塊から、培養により得られる。体外培養により無限に増殖し、生殖細胞を含むほぼ全ての組織の細胞に分化することができる。ヒトES細胞も胚盤胞の内部細胞塊から樹立されるが、マウスES細胞とは異なる性質を有することが知られている。
- 注5) サイトカイン
- 細胞間で情報を伝達する因子。多くの種類があり、細胞の分化、形態変化、遊走、成長、増殖などに影響を与える。ヒト始原生殖細胞の形成に必須なサイトカインは不明であった。
- 注6) プライム型とナイーブ型の多能性
- プライム型の多能性とは、ヒトES細胞やヒトiPS細胞に代表される細胞の有する多能性で、三胚葉[内胚葉(消化管など)、中胚葉(血液、筋肉、骨、腎臓など)、外胚葉(神経、表皮など)]系統に分化する能力は有するが、キメラ形成能を持たず、また生殖細胞分化能も低い、もしくは無いとされる。
- ナイーブ型の多能性とは、現在のところ、げっ歯類(マウス、ラット)のES細胞やiPS細胞でのみ報告されている多能性で、三胚葉系統に分化する能力は有し、キメラ形成能・生殖細胞分化能も有する多能性のこと。プライム型の多能性のヒトES細胞/iPS細胞をナイーブ型に変換しようとする研究が盛んである。
- 注7) 初期中胚葉様細胞(incipient mesoderm-like cells:iMeLCs)
- 中胚葉とは、発生初期にエピブラスト(胚体外胚葉)から誘導される3胚葉(外胚葉、中胚葉、内胚葉)の1つで、血液、筋肉、骨、腎臓等に分化する。初期中胚葉様細胞とは、エピブラストから分化した直後もしくは分化する寸前の非常に初期の中胚葉系統の細胞の性質を有すると考えられる細胞のことで、本研究で、そうした細胞がヒト生殖細胞の前駆細胞となることが示唆された。
- 注8) カニクイザル
- 哺乳綱霊長目オナガザル科マカク属に分類されるサル。ヒトを含む霊長目発生機構のモデル生物として使用される。ヒトの初期発生や生殖細胞に関わる新たな知見が得られると期待される。
- 注9) 国の指針
- 「ヒトES細胞の分配及び使用に関する指針」「ヒトiPS細胞又はヒト組織幹細胞からの生殖細胞の作成を行う研究に関する指針」
- 注10) エピブラスト様細胞(Epiblast-like cells:EpiLCs)
- エピブラスト(胚体外胚葉)とは、胚盤胞の内部細胞塊から分化する細胞で、全ての体細胞と生殖細胞の起源になる一層の上皮細胞のこと。マウスES細胞やマウスiPS細胞を特定の培養条件で分化させると、エピブラストによく似た特徴を持つエピブラスト様細胞(Epiblast-like cells:EpiLCs)へと分化する。体外培養系において、マウス始原生殖細胞様細胞へと分化する能力がある。ヒトではエピブラスト様細胞の作成例は報告されておらず、ヒトES細胞やヒトiPS細胞は、エピブラスト様細胞より分化した細胞の特徴を有するのではと考えられていた。
- 注11) エピゲノムリプログラミング
- 個体を構成するほとんど全ての細胞が有する遺伝情報は受精卵に由来し、同一であるにもかかわらず、細胞ごとに遺伝子発現パターンは異なり、多彩な機能を発揮する。これは細胞ごとに、使用される遺伝子、使用されない遺伝子が適切な化学修飾により発現を制御されているからで、そのような化学修飾のことをエピジェネティック修飾という。エピジェネティック修飾が全遺伝子レベルでいっせいに変換される現象をエピゲノムリプログラミングと呼び、生殖細胞の発生過程で起こる。
- 注12) ゲノム編集技術
- 遺伝情報を、細菌由来の遺伝子分解・切断酵素などを用いて、任意に改変する技術のこと。近年爆発的に発展しつつある。
- 注13) TFAP2C
- TFAP2C は、マウス始原生殖細胞の形成に必須な遺伝子の1つ。始原生殖細胞における機能は不明な点が多い。ヒト始原生殖細胞の発生におけるTFAP2C の機能も不明である。
- 注14) 転写制御因子
- 遺伝子が機能するには、DNAの情報がまずメッセンジャーRNAへと転写され、その後タンパク質へと翻訳される必要がある。このうち転写を制御するのが転写制御因子である。生物の発生過程では様々な転写制御因子によってゲノムのどの情報を利用するかが規定され、様々な細胞が生み出される。細胞内では多数の転写制御因子が機能しているが、細胞の種類に応じて、特徴的な転写制御因子は異なる。
- 注15) 細胞表面マーカー
- 細胞の形質膜表面に提示される膜タンパク質で、細胞外から直接認識されるので、細胞を生かしたまま標識し、細胞を選別する目的などに頻用される。今回はINTEGLIN・6とEpCAMと呼ばれる2種類のマーカーを用いた。
<論文タイトル>
“Robust In Vitro Induction of Human Germ Cell Fate from Pluripotent Stem Cells”
(多能性幹細胞からのヒト生殖細胞運命の試験管内誘導)
佐々木 恒太郎、横林 しほり、中村 友紀、岡本 郁弘、薮田 幸宏、栗本 一基、大田 浩、守時 良演、岩谷 千鶴、土屋 英明、中村 紳一郎、関口 清俊、佐久間 哲史、山本 卓、森 崇英、Knut Woltjen、中川 誠人、山本 拓也、高橋 和利、山中 伸弥、斎藤 通紀
doi :10.1016/j.stem.2015.06.014
<お問い合わせ先>
<研究に関すること>
京都大学 大学院医学研究科 生体構造医学講座 機能微細形態学
教授 斎藤 通紀(サイトウ ミチノリ)
Tel:075-753-4335(直通) Fax:075-751-7286
E-mail:
<JST事業に関すること>
科学技術振興機構 研究プロジェクト推進部
大山 健志(オオヤマ タケシ)
Tel:03-3512-3528 Fax:03-3222-2068
E-mail:
<報道担当>
京都大学 企画・情報部 広報課
Tel:075-753-2071(直通) Fax:075-753-2094
E-mail:
科学技術振興機構 広報課
Tel:03-5214-8404 Fax:03-5214-8432
E-mail: