理化学研究所(理研) 長田抗生物質研究室の中野 雄司 専任研究員、嶋田 勢津子 特別研究員(現:理研 環境資源科学研究センター 合成ゲノミクスチーム 研究員)、長田 裕之 主任研究員と、東京大学の浅見 忠男 教授(理研 長田抗生物質研究室 客員主管研究員)らの共同研究グループ※は、植物の成長促進に関わる植物ステロイドホルモン注1)「ブラシノステロイド注2)」のシグナル伝達を抑制するBSS1タンパク質が、細胞内で「集合と拡散」を行うことにより植物の草丈を制御する新しい仕組みを発見しました。
ブラシノステロイドは、植物の成長に重要な役割を果たすことが知られていますが、非常に高価なために農業や植物バイオマスの増産には利用されていませんでした。また、ブラシノステロイドが、植物の中でどのようなシグナル伝達を行い、機能しているかは明らかにされていませんでした。
共同研究グループは、ブラシノステロイドの生合成を自在に制御できる阻害剤「ブラシナゾール(Brz)注3)」を実験植物シロイヌナズナに添加し、ケミカルバイオロジー法注4)による実験を行いました。その結果、ブラシノステロイドのシグナル伝達を抑制的に制御するBSS1タンパク質を発見しました。BSS1タンパク質の細胞内での動きを調べた結果、Brzによりブラシノステロイドを欠損した状態ではBSS1タンパク質は「集合」して大きなタンパク質の塊を形成して植物の茎の伸長を抑制しました。一方、ブラシノステロイドを添加した場合は、BSS1タンパク質は「拡散」して塊は消失し、茎の伸長が促進されました。同時に、ブラシノステロイドのマスター転写因子BIL1タンパク質注5)が、BSS1タンパク質の「集合」によって捕捉され、「拡散」によって開放されることで、BIL1タンパク質の細胞質から細胞核への移行が制御されるメカニズムも明らかになりました。これらの発見により、植物の草丈を自在に制御できる技術の開発が進み、植物バイオマス増産や植物への二酸化炭素固定促進のための基盤技術の開発につながると期待できます。
本研究は、科学技術振興機構(JST) 戦略的創造研究推進事業(CREST)の一環として行われ、成果は、2015年2月6日(現地時間)に米国の科学雑誌『The Plant Cell 』オンライン版に掲載されます。
本研究は、科学技術振興機構(JST) 戦略的創造研究推進事業(CREST)「二酸化炭素資源化を目指した植物の物質生産力強化と生産物活用のための基盤技術の創出」研究領域(研究総括:磯貝 彰(奈良先端科学技術大学院大学 名誉教授))における研究課題「植物ホルモン間クロストークと化学・生物学的制御技術を利用したバイオマス高生産性植物の開発」(研究代表者:浅見 忠男、共同研究者:中野 雄司)の一環として行われました。
<背景>
植物ステロイドホルモンであるブラシノステロイドは、植物の成長に重要な役割を果たしています。しかし、ブラシノステロイドは非常に高価で、農業や植物バイオマス増産には利用されていませんでした。また、ブラシノステロイドの生合成に関する研究は大部分が明らかになってきていますが、植物の中でどのように機能しているのかは明らかにされておらず、基礎研究と応用研究の両面で、その解明が待たれています。
2002年に、中野 雄司 専任研究員、浅見 忠男 教授らは、ブラシノステロイドの生合成を自在に制御できる阻害剤「ブラシナゾール(Brz)」を用いたケミカルバイオロジー研究によって、新しい遺伝子BIL1 を発見しています注)。その後、BIL1 に由来するタンパク質「BIL1タンパク質」は、実験植物シロイヌナズナのゲノム上の約30,000種類ある遺伝子の内、約3,000種類にも及ぶ多種類の遺伝子の発現調節を行う転写因子であることが明らかにされ、現在では、BIL1タンパク質が、ブラシノステロイドのシグナル伝達のマスター転写因子(遺伝子の発現を連鎖的に引き起こす因子)であると考えられています。
BIL1タンパク質は、その転写制御対象の広さから考えられる重要性に加えて、ブラシノステロイドを添加すると細胞質から細胞核内に移行する、という興味深い動態(ダイナミクス)を示すことが知られています。しかし、細胞核移行の制御機構については、詳細な仕組みは明らかにされていません。今回、共同研究グループは、草丈制御のメカニズムの解明と、草丈制御におけるBIL1タンパク質の細胞核への移行の制御メカニズムの解明に取り組みました。
- 注) 2002年4月19日プレスリリース
- 「植物の成長を促進させる新しい因子を世界で初めて発見」
- http://www.riken.jp/~/media/riken/pr/press/2002/20020419_1/20020419_1.pdf
<研究手法と成果>
共同研究グループは、ブラシノステロイド生合成の阻害剤Brzを実験植物シロイヌナズナに添加し、ケミカルバイオロジー法による実験を行いました。Brz処理を行った野生型株を暗所で育てると、Brz無処理の野生株よりも胚軸(茎になる組織)の長さが短くなります。今回、この個体よりも胚軸の長さがさらに短い「変異体bss1(brz-sensitive short hypocotyl1)」を発見しました。このbss1 変異体の遺伝子発現などを調べた結果、ブラシノステロイドのシグナル伝達経路が抑制的に制御されていることが予測されました(図1)。
bss1 変異体の原因遺伝子を探索したところ、タンパク質同士の相互作用をするアミノ酸配列「Ankyrin リピートドメイン注6)」を持つ遺伝子が、変異体内で高発現しており、この遺伝子をBSS1 と名付けました。
次に、BSS1 に由来するBSS1タンパク質が野生型よりも高発現している変異体(BSS1 活性化型変異体)とBSS1タンパク質を高発現するように遺伝子組み換えした個体(BSS1 活性化型形質転換体)を比較しました。その結果、どちらも茎がほとんど見られず、bss1 変異体の原因がBSS1タンパク質の高発現であることが分かりました(図2)。さらに、BSS1 を欠損させBSS1タンパク質を作れないようにした個体(BSS1 欠損型変異体)は、野生型よりも草丈が長くなりました(図3)。
続いて、BSS1タンパク質の細胞内での動きを、BSS1タンパク質と緑色蛍光タンパク質GFP(Green Fluorescent Protein)との融合タンパク質(BSS1:GFP)を使って調べました。その結果、BSS1タンパク質は主に細胞質でドッド状(点状)の明るい蛍光シグナルで観察されました。このドット状の蛍光シグナルは、Brz処理を行うと数が増え、逆に、ブラシノステロイドを添加すると消失し、細胞質に薄く広がった蛍光シグナルが観察されました。
さらに、詳しい解析の結果、蛍光シグナルが、ドット状で見られるときは、BSS1:GFP同士が「集合」し、結合しあって大きなタンパク質の塊を形成していること、細胞質に広がって観察されるときは、BSS1:GFPは「拡散」して、単独のタンパク質として存在していることが分かりました(図4)。ブラシノステロイドの増減に伴って、タンパク質が集合と拡散という動態を示すことは、他の植物ホルモンなどの生理活性物質では見られることの少ない非常に珍しい現象です。
BSS1タンパク質が細胞質と細胞核に存在することから、同じように細胞質と細胞核に存在するBIL1タンパク質との関わりを調べました。
はじめに、酵母のツーハイブリッド法注7)や、植物体内の免疫沈降実験注8)でBIL1タンパク質とBSS1タンパク質が互いに結合する性質を持つことが明らかになりました。次に、BSS1タンパク質の発現を欠損させた個体(BSS1 欠損型変異体)では、BIL1タンパク質の細胞核内での局在量が増加すること、BSS1タンパク質の発現量を増加させた個体(BSS1 活性化型形質転換体)では、BIL1タンパク質の細胞核内での存在量が減少することが明らかになりました(図5)。
これらの結果は、BSS1タンパク質が、BIL1タンパク質と結合することによって、BIL1タンパク質を細胞質に引き止めていることを示し、BIL1タンパク質の細胞核移行においてBSS1タンパク質が抑制的な役割を果たしていることが明らかになりました。
これらの結果を整理したBSS1タンパク質の機能についてのモデル図を図6に示します。Brzによりブラシノステロイドを欠損させると、BSS1タンパク質は「集合」して、大きなタンパク質の塊(複合体)となり、BIL1タンパク質をその塊に捕捉します。捕捉されたBIL1タンパク質は細胞核へ移行できなくなり、その下流で転写制御を受けるはずだった遺伝子の発現は生じなくなります。その結果、植物の茎の伸長は低下し、草丈は短くなります。
一方、ブラシノステロイドを添加すると、BSS1タンパク質の塊は「拡散」して、細胞質に広がって存在するようになります。単体となったBSS1タンパク質は、BIL1タンパク質を捕捉する力が弱まり、開放されたBIL1タンパク質は細胞質から細胞核へ移行するようになります。細胞核内では、BIL1タンパク質による遺伝子発現が活性化され、その結果、植物の茎の伸長は促進し、草丈は長くなります。
これらの結果から、植物の成長(草丈の伸長)の制御が、BSS1タンパク質の「集合」と「拡散」というタンパク質の動態によってコントロールされているという新しい制御メカニズムを明らかにしました。
<今後の期待>
本研究によって得られたBSS1 の遺伝子情報と知見を活用することによって、植物の草丈を自在に制御する技術開発が可能になると考えられます。
植物は、大気中の二酸化炭素を細胞壁など自らの体に固定する能力において、地球温暖化対策の重要な鍵となります。また、固定された炭素は、植物バイオマスの原料となり、クリーンなバイオ燃料の源にもなります。植物の茎は、葉と並んで、植物バイオマスの量を決める重要な器官です。
また、途上国などで深刻化している食糧不足の問題に対しては、根源的な作物生産量の増大が最も近い解決策であると考えられます。穀物生産量の増加には、最適な長さに植物の草丈を制御する技術が必須であり、その改良は非常に重要であると考えられています。
現時点での知見は、実験植物シロイヌナズナで得られたものですが、BSS1 遺伝子は、植物種を越えて、イネやトウモロコシ、サトウキビなどにも広く保存されている遺伝子です。これらの実用作物におけるBSS1遺伝子へと研究を発展させることによって、地球環境の保護や食糧増産などに貢献すると期待できます。
※ 共同研究グループ
| 理化学研究所 |
| | 長田抗生物質研究室 |
| | 専任研究員 | 中野 雄司(なかの たけし) |
| 特別研究員 | 嶋田 勢津子(しまだ せつこ)
(現:環境資源科学研究センター バイオマス工学連携部門 合成ゲノミクスチーム 研究員) |
大学院生リサーチ
・アソシエイト | 小松 知之(こまつ ともゆき)(東京農工大学 大学院博士課程)
(現:日本たばこ産業(株)) |
| 特別研究員 | 山上 あゆみ(やまがみ あゆみ) |
| 主任研究員 | 長田 裕之(おさだ ひろゆき) |
| | 環境資源科学研究センター バイオマス工学連携部門 合成ゲノミクス研究チーム |
| | チームリーダー | 松井 南(まつい みなみ) |
| 研究員 | 中澤 美紀(なかざわ みき) |
| 東京大学 大学院農学生命科学研究科 応用生命化学専攻 生物制御化学研究室 |
| | 教授 | 浅見 忠男(あさみ ただお)
(理研 長田抗生物質研究室 客員主管研究員) |
| 東京農工大学 大学院農学府 生物制御科学専攻 生物制御化学研究室 |
| | 教授 | 夏目 雅裕(なつめ まさひろ) |
| 准教授 | 川出 洋(かわいで ひろし) |
<参考図>
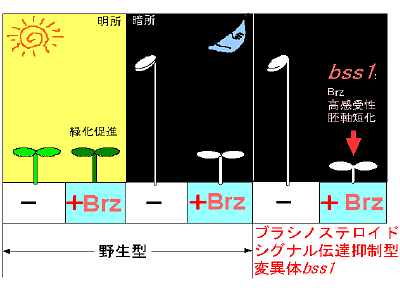
図1 bss1 変異体の探索
Brz処理を行った野生型株を暗所で育てると、Brz無処理の野生株よりも胚軸の長さが短くなる。bss1 変異体は、この個体よりもさらに胚軸が短い。これは、bss1 変異体の中で、ブラシノステロイドのシグナル伝達の抑制因子が高発現している可能性を示している。

図2 シロイヌナズナのBSS1 活性化型変異体とBSS1活性化型形質転換体
BSS1 活性化型変異体とBSS1 活性化型形質転換体は、共に、茎が極端に短くなる。BSS1タンパク質を野生型において高発現するように遺伝子組み換えした植物が、bss1 変異体と同じ形態を示したことは、BSS1タンパク質の高発現が、bss1 変異体の原因であることを証明したことになる。

図3 シロイヌナズナの野生型(左)と BSS1欠損型変異体(右)
BSS1 欠損型変異体は、野生型よりも草丈が長くなる。
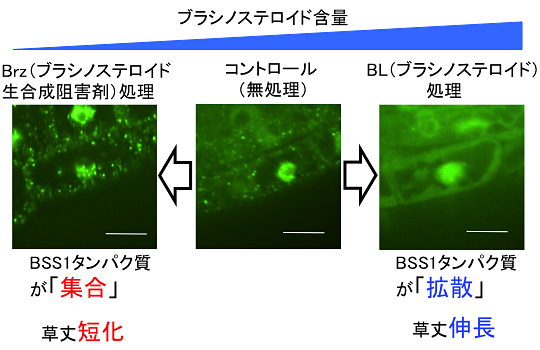
図4 BSS1:GFPの顕微鏡観察像
ブラシノステロイド欠損(Brz処理)で見られるBSS1タンパク質の「集合」と、ブラシノステロイド添加(BL処理)で見られるBSS1タンパク質の「拡散」。
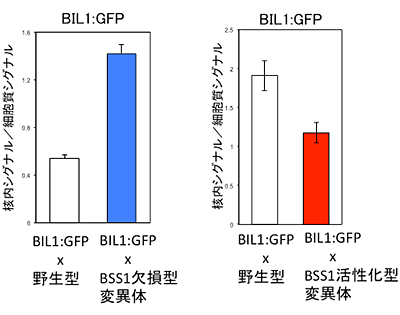
図5 BSS1タンパク質欠損/活性化による
BIL1-GFPタンパク質の細胞核局在への影響
- 左:BSS1タンパク質欠損によりBIL1-GFPタンパク質の細胞核局在が増加する。
- 右:BSS1タンパク質活性化によりBIL1-GFPタンパク質の細胞核局在が減少する。
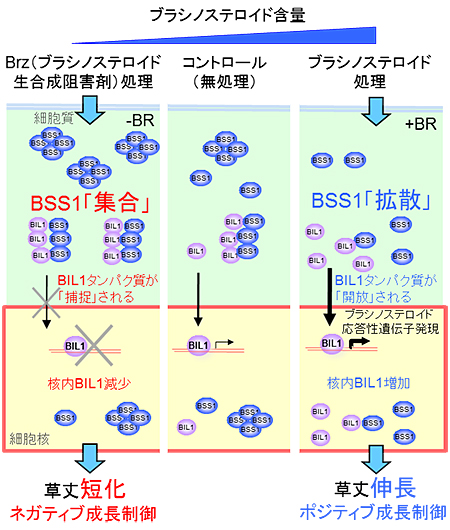
図6 BSS1タンパク質の機能発現のモデル
- 左:Brzによりブラシノステロイドを欠損させると、BSS1タンパク質は「集合」して、大きなタンパク質の塊(複合体)となり、BIL1タンパク質を捕捉する。捕捉されたBIL1タンパク質は細胞核へ移行できなくなり、その下流で転写制御を受けるはずだった遺伝子の発現は生じなくなる。その結果、植物の茎の伸長は低下し、草丈は短くなる。
- 右:ブラシノステロイド添加において、BSS1タンパク質の塊は「拡散」して、細胞質に広がって存在するようになる。単体となったBSS1タンパク質は、BIL1タンパク質を捕捉する力が弱まり、開放されたBIL1タンパク質は細胞質から細胞核へ移行するようになる。細胞核内では、BIL1タンパク質による遺伝子発現が活性化され、その結果、植物の茎の伸長は促進し、草丈は長くなる。
<用語解説>
- 注1) ステロイドホルモン
- 多細胞生物が種を越えて広く持っている生理活性化合物。ステロイド骨格と呼ばれる4員環骨格を持ち、ほ乳類では筋肉などを作る男性ホルモンのテストステロン、妊娠の維持に関わる女性ホルモンのプロゲステロン、昆虫の脱皮に関係するエクダイソンが知られる。植物でもステロイドホルモンは存在し、ブラシノステロイドが植物の成長や葉緑体の発達を制御していることが明らかになりつつある。
- 注2) ブラシノステロイド
- 1979年に米国農務省の研究グループがアブラナの花粉から発見し、化学構造を決定した。アブラナの学名であるBrassica(ブラシカ)が名前の由来で、7種類(オーキシン、サイトカイニン、ジベレリン、アブシジン酸、エチレン、ブラシノステロイド、ストリゴラクトン)ある植物ホルモンの1つと定義される。しかし、ほかの植物ホルモンが植物に固有の化合物であり、また、葉緑体内で生合成される酵素反応の段階があることや、植物に限定的な生理活性が知られていることに対して、ブラシノステロイドは生合成経路のすべてが細胞質で行われること、動物や昆虫にも類縁化合物であるステロイドホルモン類が存在し、各々の生物種においても生理活性を持つなど「生物種間で普遍的」である点が特徴。植物に対しては、細胞伸長や細胞分裂の促進など細胞レベルの促進的作用や、子葉の開化、胚軸の伸長、維管束の分化、緑葉の上偏成長、葉柄の伸長、茎の伸長など器官レベルの制御作用、また、ストレス耐性の付与(耐冷、耐塩、耐乾燥)や植物病害抵抗性の促進(植物自然免疫の活性化)など、さまざまな生理作用を示す。
- 注3) ブラシナゾール(Brz)
- 植物体のブラシノステロイド生合成経路を阻害する化合物。1998年に理研植物機能研究室の浅見忠男博士(現:東京大学教授)らが創製した。Brzを植物に処理することにより、生合成欠損変異体と同じような形、大きさを植物に与えることができる。従来の一般的な方法、つまり遺伝子を変異させてブラシノステロイド欠損状態にした植物を入手する方法では、成長の最初から最後まで、決まった量のブラシノステロイド欠損状態しか植物に与えることができない。しかし、このBrzを使用することにより、すべての植物のあらゆる成長時期や部位で、しかも量の多少を調節して、ブラシノステロイド欠損状態をもたらすことができる。この手法により、植物成長におけるブラシノステロイドの生理作用をより詳細に明らかにすることが可能となった。
- 注4) ケミカルバイオロジー法
- 化学物質(ケミカル)の力により、生命の仕組み(バイオロジー)を明らかにすることを試みる研究手法。生物の形態やタンパク質の働きに対して生理活性を示す小分子化合物を人工的な有機合成により創製し、それを利用して標的タンパク質分子の同定やタンパク質の機能解明を目指す手法などをいう。有機合成化学を出発点とすることから、タンパク質の機能解明により生物自身が体内で合成する天然物の働きを明らかにすることを主とする通常の「生化学的」手法とは区別して考えることが多い。
- 注5) BIL1タンパク質
- 2002年、理研植物機能研究室の中野 雄司 研究員(現:長田抗生物質研究室 専任研究員)、浅見 忠男 副主任研究員(現:東京大学 教授)らの研究チームがブラシノステロイド生合成阻害剤Brzを用いたケミカルバイオロジー研究で単離した突然変異体bil1(Brz-insensitive-long hypocotyl1) の原因遺伝子として同定した遺伝子BILに由来するタンパク質。同時期に、米国ソーク研究所でも独立してbzr1(Brz-resistant1) の突然変異体の原因遺伝子BZR1が同定された。BIL1 とBZR1 は同一遺伝子である。BIL1(BZR1)タンパク質は、ブラシノステロイド刺激によって細胞質から核に移行し、ブラシノステロイド刺激を核内に伝える働きをしている。その後、プロテアソーム系によって分解され、ブラシノステロイド刺激は比較的短時間でリセットされる。
- 注6) Ankyrinリピートドメイン
- ループ構造を挟んだ2つのアルファへリックス構造を構成する33アミノ酸からなるアミノ酸モチーフが10数回繰り返される特徴的な立体構造を持つドメイン(領域)。真核生物の植物から動物までの多種生物種のタンパク質に広く保存されており、少数ながら原核生物にも存在することも知られている。ドメインの分子機能としては、タンパク質-タンパク質間の相互作用を担う役割が主に知られる。また、このドメインを持つタンパク質には、タンパク質間相互作用が機能発現に必須なシグナル伝達タンパク質群を始めとして、転写因子、イオン輸送体タンパク質などがある。
- 注7) ツーハイブリッド法
- 植物の2種類のタンパク質が直接結合するか否かについて、酵母の細胞系を用いて解析する実験手法。
- 注8) 免疫沈降実験
- 植物の2種類のタンパク質が直接結合するか否かについて、それぞれのタンパク質を同時に発現させた植物細胞から精製したタンパク質を用いて解析する実験手法。
<論文情報>
| タイトル |
“Formation and Dissociation of the BSS1 Protein Complex Regulates Plant Development via Brassinosteroid Signaling” |
| 著者名 |
Setsuko Shimada, Tomoyuki Komatsu, Ayumi Yamagami, Miki Nakazawa, Minami Matsui, Hiroshi Kawaide, Masahiro Natsume, Hiroyuki Osada, Tadao Asami, and Takeshi Nakano |
| 雑誌名 |
The Plant Cell |
| doi |
10.1105/tpc.114.131508 |
<お問い合わせ先>
<研究に関すること>
中野 雄司(ナカノ タケシ)
理化学研究所 長田抗生物質研究室 専任研究員
Tel:048-467-9043 Fax:048-467-4389
E-mail:
嶋田 勢津子(シマダ セツコ)
理化学研究所 長田抗生物質研究室 特別研究員
(現:理研 環境資源科学研究センター バイオマス工学連携部門 合成ゲノミクスチーム 研究員)
長田 裕之(オサダ ヒロユキ)
理化学研究所 長田抗生物質研究室 主任研究員
浅見 忠男(アサミ タダオ)
東京大学 大学院農学生命科学研究科 応用生命化学専攻 生物制御化学研究室 教授
(理研 長田抗生物質研究室 客員主管研究員)
<JST事業に関すること>
科学技術振興機構 戦略研究推進部
Tel:03-3512-3531 Fax:03-3222-2066
E-mail:
<報道担当>
理化学研究所 広報室 報道担当
Tel:048-467-9272 Fax:048-462-4715
E-mail:
科学技術振興機構 広報課
Tel:03-5214-8404 Fax:03-5214-8432
E-mail: