ポイント
- 植物ではこれまで困難であった組織単位での「時計遺伝子」発現の定量解析に初めて成功。
- 維管束の時計遺伝子の機能を阻害するだけで花の咲くタイミングを遅らせることができた。
- 植物組織の体内時計機能は、植物の精密な生長調節法開発のターゲットとして期待。
JST 戦略的創造研究推進事業において、京都大学の遠藤 求 助教らは、植物組織の遺伝子発現をモニタリングする新規解析手法を開発し、時計遺伝子の発現を組織レベルで定量的に測定することに成功しました。
動物では、脳に存在する体内時計注1)とその他の臓器に存在する体内時計の機能が異なることが知られていました。植物でも、動物のような体内時計の機能分担が組織レベルで存在する可能性は指摘されていましたが、組織単離に時間がかかるため、時々刻々と発現量が変化する体内時計に関わる時計遺伝子の発現を、定量的・経時的に測定することは困難でした。
本研究グループは、各組織での時計遺伝子の発現を迅速に測定するために、組織単離時間を従来法の1/3以下に短縮しました。また、時計遺伝子の発現を非侵襲で測定できる「TSLA法」を世界で初めて開発し、維管束に存在する時計遺伝子の性質が他の組織と大きく異なり、隣接する葉肉組織の時計遺伝子の発現に影響を与えていることを明らかにしました。さらに、維管束の時計機能を阻害するだけで植物の花の咲くタイミングを遅らせることにも成功しました。
本研究の結果は、植物がどのように時間を測りその情報を個体レベルで統合しているのかを解明する手がかりになるだけでなく、植物の組織単位の時計機能をターゲットにすることで植物の精密な生長調節法の開発が期待されます。
本研究成果は、2014年10月29日(英国時間)に英国科学誌「Nature」のオンライン速報版で公開されます。
本成果は、以下の事業・研究領域・研究課題によって得られました。
戦略的創造研究推進事業 個人型研究(さきがけ)
| 研究領域 |
「細胞機能の構成的な理解と制御」
(研究総括:上田 泰己 東京大学 大学院医学系研究科 教授) |
| 研究課題名 |
「構成的アプローチによる植物の生物時計の組織特異的な役割の解明」 |
| 研究者 |
遠藤 求(京都大学 大学院生命科学研究科 助教) |
| 研究実施場所 |
京都大学 農学・生命科学研究棟721 |
| 研究期間 |
平成23年12月~平成26年11月 |
<研究の背景と経緯>
これまで植物の体内時計は、個々の細胞が独自に時間を測っており、どの組織の体内時計も同じ機能を持っていると漠然と考えられていたため、植物個体全体を用いた解析が主流であり(図1)、植物組織ごとの体内時計機能や組織・細胞間での体内時間情報のやりとりについてはほとんど解析されてきませんでした。
動物では、外科的な手術により脳など特定の組織の一部を切除・移植するといった実験や培養細胞を用いた実験から、各組織の体内時計の役割、特に脳にある体内時計の重要性が明らかにされてきました。しかし、植物は脳に対応する明確な中枢を持たず、機能を維持したまま組織を培養することも困難でした。また植物では、動物とは異なり細胞壁によって細胞同士が固く接着しているため組織単離に非常に時間がかかり、時間の経過とともに発現量が変化する、時計遺伝子の発現を定量的・経時的に測定することは困難でした。植物の体内時計の組織特異的注2)な解析を進展させるために、こうした技術的な課題を解決することが強く望まれていました。
<研究の内容>
遠藤助教らはまず、これまで組織レベルでの体内時計の解析を難しいものにしていた組織単離法の改良に取り組みました。これまでは、目的組織でのみ蛍光たんぱく質で標識した植物から、蛍光を指標として組織を単離していたため、単離のために約1時間30分~4時間30分ほどかかっていました。そのため、操作中に時計遺伝子の発現量が変化してしまい、体内時計を定量的に測定することは困難でした。今回、超音波処理と酵素処理を組み合わせることで、シロイヌナズナ注3)からの組織単離にかかる時間を30分以内に短縮することに成功しました。これにより、単離操作中の遺伝子発現量の変化をほとんど気にする必要がなくなり、初めて定量的に各組織の体内時計を解析することが可能になりました(図2)。この方法を用いて、シロイヌナズナに光を当て始めてからの葉全体、葉肉組織と維管束組織の、時計遺伝子の発現を定量的に測定したところ、発現量や発現リズム、標的遺伝子などが、葉全体と葉肉の発現様式はよく似ているのに対し、維管束はそのどちらとも大きく異なっていることがわかりました(図3)。こうしたことから、植物でも動物と同様に体内時計システムは組織ごとに異なっていることが考えられました。
さらに、維管束で時計の働きを阻害すると葉肉の体内時計の働きも阻害される一方で、葉肉の時計の働きを阻害しても維管束の時計の働きには影響がないこともわかりました(図4)。こうした時計制御の非対称性は動物でも見られており、植物の体内時計システムは動物と同様に階層構造を持っていることも明らかになりました。
このことを別のアプローチから確認するために、組織レベルでの遺伝子発現を非侵襲で測定するための新しい方法としてTissue-specific luciferase assay(TSLA)法を世界で初めて開発しました(図5)
。従来法では、全ての組織で発光リズムが見られるため、特定の組織における発光量を定量することは困難でしたが、この方法では、ルシフェラーゼ注4)という酵素の遺伝子を2つに分け、組織特異的プロモーター注5)および時計プロモーターによって発現させ、2つの発現が重なる時間・空間のみでルシフェラーゼを再構成させることで、特定の組織における時計遺伝子の発現を発光リズムとしてとらえることができます。TSLA法を用いた解析から、同じ時計遺伝子でも維管束と葉全体では異なる遺伝子発現リズムが見られることからも、葉肉と維管束の時計システムが異なっていることが支持されました(図6)
。
さらに、こうした維管束の体内時計はフロリゲン注6)と呼ばれる花成ホルモンの産生を通じて、個体全体の生理応答を制御していることも示され、植物の体内時計において維管束が非常に重要であることが示されました(図7)
。
<今後の展開>
体内時計は多くの遺伝子発現の制御に関わっていますので、花成や細胞伸長など体内時計によって制御されている生理応答の解析も、組織レベルで行っていく必要があることがわかりました。本研究で開発した手法を用いることで、こうした組織レベルでの解析が大きく進むことが期待されます。
また、維管束の時計機能を阻害するだけで植物の花の咲くタイミングを遅らせることができたことから、体内時計は植物の生長調節法開発の新たなターゲットになる可能性が期待されます。
<参考図>
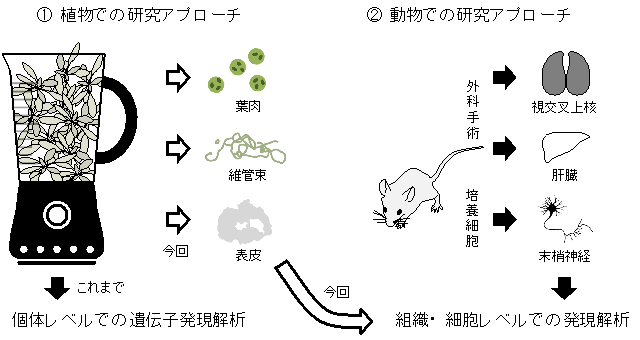
図1 動物と植物における体内時計研究アプローチの違い
- ① これまで、植物の体内時計研究は個体全体をすりつぶして遺伝子発現解析を行っていたので、体内時計の組織レベルでの振る舞いは解析できなかった。
- ② 動物細胞では外科手術で組織を摘出する方法や、培養細胞を使用することで体内時計の組織レベルでの振る舞いを解析することができる。
今回、高効率・短時間で植物の組織を単離する方法を開発することで、初めて組織レベルでの発現解析を実現できた(詳細は図2参照)。
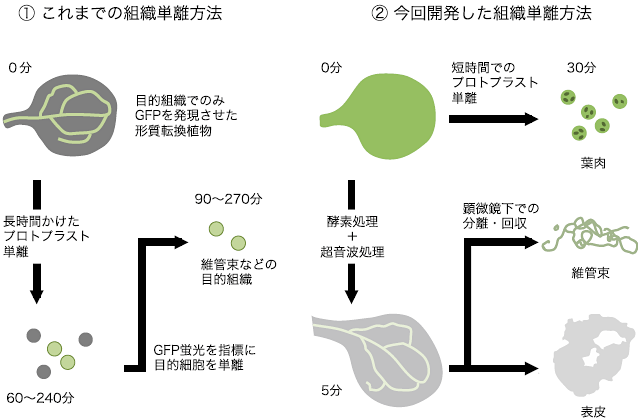
図2 植物におけるこれまでの組織単離法と今回開発した組織単離法の違い
- ① これまでの方法では、緑色蛍光たんぱく質GFPを植物の組織特異的に発現させ、プロトプラスト(酵素処理により細胞壁を取り除いた植物細胞)を単離し、さらにGFPを指標に目的の組織を単離していた。この方法では単離した組織の純度は高いが、時間がかかってしまうため(1時間30分~4時間30分程度)、時間とともに発現が変化する体内時計の解析には不向きであった。
- ② 今回開発した方法では、葉を短時間酵素処理することで、葉肉だけをプロトプラストとして単離した後、葉に酵素処理と超音波処理を組み合わせて行うことで、葉肉を破砕し表皮と維管束だけを効率よく単離することができる。これにより、組織単離に要する時間を30分以内に短縮することができ、時間経過に伴う発現量変化をほとんど気にすることなく組織レベルでの遺伝子発現解析を行うことができる。
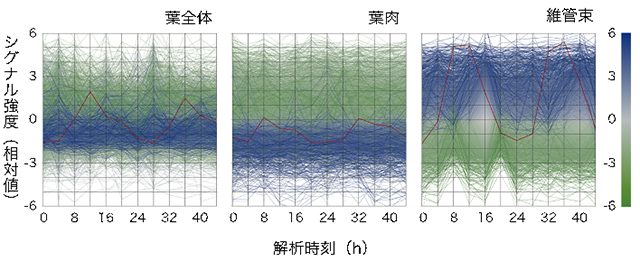
図3 維管束は、葉全体や葉肉とは全く異なる発現様式を持つ
24時間のうち、16時間は光が当たる環境(明期)、8時間は光が当たらない環境(暗期)で生育したシロイヌナズナの葉全体、葉肉と維管束の遺伝子発現を解析した。体内時計によって制御されている遺伝子のうち、維管束における遺伝子発現量の平均を0とし、高発現の遺伝子を青線で、低発現の遺伝子を緑線で塗り分けた。横軸は明期開始を0としたときの経過時間を示している。
葉全体と葉肉の発現様式はよく似ているが、維管束はそのどちらとも大きく異なり、葉全体や葉肉で発現の低い遺伝子(青線)が高発現している。維管束で特異的に発現していることが明らかとなった時計遺伝子ELF4の発現を赤い線で例示した。
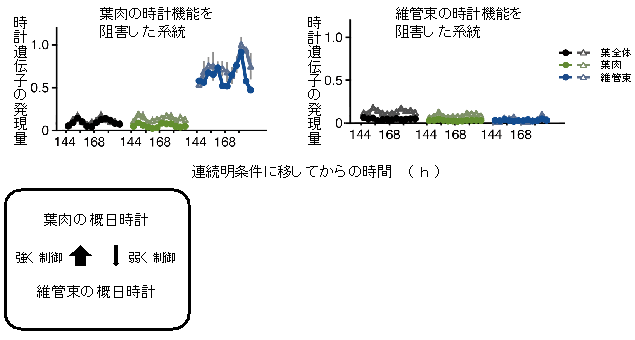
図4 葉肉と維管束の体内時計は非対称的な制御関係を持つ
- (図左) 遺伝子操作により葉肉の時計機能を阻害したシロイヌナズナでは、葉全体(黒線)と葉肉(緑線)での時計遺伝子の発現は低下するが、維管束(青線)でのリズムは消失しない
- (図右) 一方、遺伝子操作により維管束の時計機能を阻害したシロイヌナズナでは、維管束(青線)だけでなく葉全体(黒線)や葉肉(緑線)の時計遺伝子の発現まで消失する。
このことは、維管束の体内時計が葉肉の体内時計に対して支配的であることを示している。
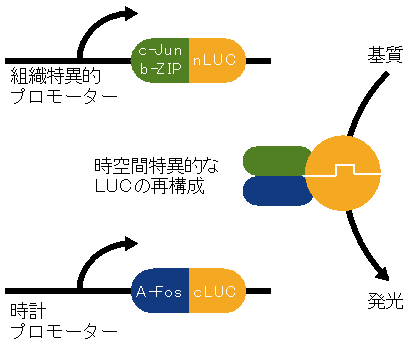
図5 Tissue-specific luciferase assay(TSLA)法の原理
ルシフェラーゼ(LUC)を断片化したもの(nLUCとcLUC)を、それぞれJun遺伝子とFos遺伝子の部分断片を改変したもの(c-Jun b-ZIPとA-Fos)と融合させる。その後、一方を目的の組織特異的プロモーター、もう一方を時計プロモーターで発現させる。たんぱく質に翻訳された両者は、JunとFosを介して特異的に結合し(Junたんぱく質とFosたんぱく質は結合することが知られている)、断片化されていたルシフェラーゼが再構築される。2つの発現が重なる時間・空間のみでルシフェラーゼを再構成させることで、目的の組織で目的の時計遺伝子の発現が発光リズムとして検出できる。
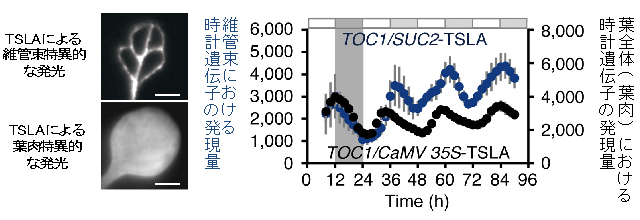
図6 TSLA法による組織特異的な時計遺伝子発現の解析
- (図左上) TSLA法で維管束特異的に時計遺伝子を発光させた。
- (図左下) TSLA法で葉肉特異的に時計遺伝子を発光させた。
- (図右)時計遺伝子TOC1の発現量を示す。TSLA法では発光が特定の組織に限定されるので、組織特異的な発光リズムを定量的に計測することができる。測定の結果、葉肉(黒丸)と維管束(青丸)では同じ遺伝子でも異なるリズムが見られたことから、葉肉と維管束の体内時計は異なると考えられる。横軸は連続明条件にしてからの時間を表す。

図7 維管束の体内時計機能を阻害すると遅咲き表現型を示す
24時間のうち、16時間は光が当たる環境(明期)、8時間は光が当たらない環境(暗期)で1ヶ月間生育させたシロイヌナズナ。維管束を含む組織(全体と維管束)で、遺伝子操作により体内時計機能を阻害したシロイヌナズナは、顕著に花が咲くのが遅くなる一方で、葉肉や表皮で体内時計機能を阻害しても野生型(遺伝子操作をしていないシロイヌナズナ)と同様に花が咲くことがわかる。
<用語解説>
- 注1) 体内時計
- 約24時間周期のリズムを生み出すための仕組み。概日時計。地球の自転や公転に伴う明暗周期や季節変化に対応するための仕組みとして獲得されたと考えられている。
- 注2) 組織特異的
- ここでは、ある遺伝子の機能が必ずしも発現している全ての組織では必要なく、特定の組織でのみ機能していることを指している。
- 注3) シロイヌナズナ
- 代表的なモデル植物。アブラナ科。多くの時計遺伝子がすでに同定されており、体内時計の仕組みの解明が最も進んでいる。
- 注4) ルシフェラーゼ
- ホタルなどの生物発光において、発光物質が光を放つ化学反応を触媒する作用を持つ酵素の総称。
- 注5) プロモーター(発現調整領域)
- DNA上で遺伝子をコードする領域の付近で、その遺伝子の転写開始・停止を調節する配列を含む領域のこと。
- 注6) フロリゲン
- 植物の花芽形成(花成)を誘導する植物ホルモン。
<論文タイトル>
“Tissue-specific clocks in Arabidopsis show asymmetric coupling”
(シロイヌナズナの組織特異的な概日時計は非対称的にカップリングしている)
doi: 10.1038/nature13919
<お問い合わせ先>
<研究に関すること>
遠藤 求(エンドウ モトム)
京都大学 大学院生命科学研究科 分子代謝制御学 助教
Tel:075-753-6142 Fax:075-753-6470
E-mail:
<JST事業に関すること>
松尾 浩司(マツオ コウジ)、川口 貴史(カワグチ タカフミ) 、山岸 裕司(ヤマギシ ユウジ)
科学技術振興機構 戦略研究推進部 ライフイノベーショングループ
〒102-0076 東京都千代田区五番町7 K’s五番町
Tel:03-3512-3524 Fax:03-3222-2064
E-mail:
<報道担当>
科学技術振興機構 広報課
〒102-8666 東京都千代田区四番町5番地3
Tel:03-5214-8404 Fax:03-5214-8432
E-mail: