<ポイント>
- 植物生理の解析や新しい育種技術開発のために、遺伝子組み換えに頼らない簡便な解析法や遺伝子の操作法が求められていた。
- S化オリゴを培地に添加するだけで、花粉管内の遺伝子の働きを抑えられることを発見し、狙った遺伝子ごとに異なる効果を確認。
- 遺伝子組み換えを使わない植物の遺伝子制御法として、育種など農業分野への応用に期待。
JST 課題達成型基礎研究の一環として、戦略的創造研究推進事業 ERATO型研究「東山ライブホロニクスプロジェクト」(研究総括:東山 哲也 名古屋大学 WPIトランスフォーマティブ生命分子研究所 教授)の水多 陽子 研究員(名古屋大学 大学院理学研究科)は、植物の花粉管注1)に試薬を与えるだけで特定の遺伝子の働きを抑えられることを発見しました。
被子植物では、めしべの先端に花粉が付着(受粉)すると、花粉から花粉管と呼ばれる管が伸びて、受精が行われることで種子が作られます。しかし、花粉管が正常に伸びていくにはどのような遺伝子の働きが必要なのか、その全容は明らかとなっていません。解析方法も突然変異体や遺伝子組み換え注2)を用いたものが中心であり、時間と手間が必要となります。また、農業分野では遺伝子組み換えを用いずに、収量増加などの育種目標を達成する新しい技術の開発が望まれています。
研究グループは、遺伝子の働きを阻害するホスホロチオエート化注3)アンチセンスオリゴDNA注4)(S化オリゴ)という試薬を含む培地に花粉を一緒に培養するだけで、花粉管の遺伝子を制御する方法を開発しました。花粉が物質を取り込みやすい性質を生かし、最適な条件下で培養すると、花粉にS化オリゴが自然に取り込まれます。その結果、狙った遺伝子の働きを抑え、実際に花粉管の伸長を制御できることを確認しました。今後、特定の遺伝子の働きを制御するなど、植物の受精の仕組みの解明に役立つことが期待されます。将来、めしべにS化オリゴを与え、めしべの上で花粉にS化オリゴを取り込ませることで、花粉管の伸長を妨げる遺伝子の働きを抑えて受精を成功させるなど、遺伝子組み換えを必要としない植物の交雑育種注5)や農業分野への応用が期待されます。
本研究成果は、英国の科学誌「Plant Journal」のオンライン速報版で近日中に公開されます。
本成果は、以下の事業・研究プロジェクトによって得られました。
戦略的創造研究推進事業 ERATO型研究
| 研究プロジェクト |
「東山ライブホロニクスプロジェクト」
|
| 研究総括 |
東山 哲也(名古屋大学 WPIトランスフォーマティブ生命分子研究所 教授) |
| 研究期間 |
平成22年10月~平成28年3月 |
多細胞生物の細胞が種々のシグナル分子を介して、他の細胞と情報のやり取りをすることで自身の振る舞いを決定し、生物としてのバランスを取るための仕組み「ホロニックコミュニケーション」の全容解明に向けた、多くのライブセル解析技術の創出を目指します。
<研究の背景と経緯>
被子植物では、めしべに花粉が付着(受粉)すると、花粉から花粉管と呼ばれる管が発芽し、めしべ内部へ伸長します。伸長した花粉管が生殖細胞(胚珠)に到達し、受精が行われることで種子が作られます(図1)。つまり、種子が実るためには花粉管が正常に発芽し、胚珠まで順調に伸びていく必要があります。しかし、花粉管が伸びていくためにはどのような遺伝子が必要なのか、その全容は明らかとなっていません。また、農業分野では、新しい有用な品種を作るため、さまざまな種・品種間で交雑が行われてきました。しかし、ある一部の組み合わせでは花粉管が正常に伸びずに種子が得られないなど、交雑時の問題も数多く知られています。これらの問題に対する知見が得られない理由の1つとして、花粉管伸長に関わる遺伝子を調べる方法は、突然変異体や遺伝子組み換え体を用いた解析が中心であり、研究に手間と時間がかかるといった点が挙げられます。加えて、農作物に対しては遺伝子組み換え体を利用することが困難であるという側面もあります。
アンチセンスオリゴDNA(アンチセンスオリゴ)は、遺伝子のmRNAと相補的な配列注6)を持ち、細胞内に取り込まれた後、標的とするmRNAと結合して遺伝子の働きを阻害します(図2)。アンチセンスオリゴは細胞にとって異物であるため、本来は取り込まれた後に細胞内で分解されてしまいます。しかし、アンチセンスオリゴをホスホロチオエート化(S化)することで、細胞内で分解されにくくなり、遺伝子の働きを継続的に阻害できるようになります。花粉、および花粉管はめしべの上で吸水して発芽・伸長することから、もともと物質を取り込みやすいという性質があります。この性質を利用して、S化したアンチセンスオリゴ(S化オリゴ)を花粉に取り込ませ、遺伝子の働きを抑える方法がいくつか報告されてきました。しかし、S化オリゴの取り込みに用いる試薬自体に毒性があることや、試薬が高価で手順が煩雑であること、そして応用範囲が種によって異なることなどから、より簡便で毒性がなく、さまざまな植物に応用可能な効果的な方法が求められていました。
<研究の内容>
上記の問題点を解決すべく、水多研究員らはS化オリゴによる簡便な花粉管内遺伝子の発現制御法の確立を目標とし、実験を行いました。実験材料には、年間を通じて花粉管の研究が可能で、花粉を大量に採取できるアブラナ科のシロイヌナズナを用いました。
まず、シロイヌナズナの花粉管でS化オリゴを用いた報告がなかったことから、花粉管の伸長に適した培養条件や、S化オリゴの適切な濃度の検討を行いました。条件検討の結果、S化オリゴの適切な濃度は10~20μM(マイクロモーラー)注7)であり、40μM以上では花粉管に悪影響を及ぼすことが分かりました。この結果から、シロイヌナズナの花粉管でS化オリゴを用いる際の最適な濃度範囲が明らかになりました。
また、この過程で、これまでの報告より簡便にS化オリゴを取り込ませる方法を見いだしました。これまでは、リポソームと呼ばれるリン脂質二重膜の小胞の内部にS化オリゴを包み込み、細胞へ取り込ませる方法(リポフェクション)が用いられていました(図3)。しかし、リポフェクションに用いる試薬は高価であり、また高濃度のリポフェクション試薬は細胞に毒性を持つことが知られています。これに対し、本研究では、最適な濃度でS化オリゴを加えた培地で花粉を培養するだけで、リポフェクションを用いることなくシロイヌナズナの花粉にS化オリゴを取り込ませ、遺伝子の働きを阻害することに成功しました(図3)。
続いて、花粉管の発芽と伸長に重要であることが報告されているANXUR(ANX) 遺伝子のS化オリゴを作製し、どのような効果が得られるかを調査しました。これまでの報告から、ANX 遺伝子の働きを阻害すると、花粉管は発芽した直後に先端が破裂してしまうことが知られていました。このことから、S化オリゴでANX 遺伝子の働きを阻害した場合も、花粉管の発芽や先端の形態に影響が生じることが予想されました。ANX 遺伝子に対するS化オリゴを培地に添加し、花粉を培地上に散布したところ、花粉管の発芽に影響はないものの、花粉管の長さが顕著に短くなりました(図4、図5)。また、花粉管にこぶや枝分かれなど、正常な花粉管では見られないような形態の変化が観察されました(図4)。これは、花粉管の先端が正常に伸長できなくなったためだと考えられます。
その後、他の遺伝子の働きも阻害できるかどうか確認するため、新たにCalS5 遺伝子、およびROP1 遺伝子についてS化オリゴを作製し、同様の実験を行いました。CalS5 遺伝子は花粉管内でカロース注8)と呼ばれる物質を合成する遺伝子です。CalS5 遺伝子の働きを抑えると、カロースで作られた栓状の構造物(カロース栓注8))を作れなくなり、花粉管の伸長に影響することが知られています。CalS5 遺伝子に対するS化オリゴを用いると、花粉の発芽には影響がないものの、花粉管内のカロース栓の数が大きく減少(図6)し、花粉管がとても短くなることが明らかとなりました。一方、ROP1 は花粉管の先端の形状を維持し、花粉管がまっすぐ伸長していくために重要な遺伝子です。ROP1 遺伝子に対するS化オリゴを用いると、花粉管が蛇行し、先端が波打つなどの形態変化が観察されました(図4)。
次に、S化オリゴによってどのように遺伝子の働きが抑えられるのか、そのメカニズムを調べるため、S化オリゴで狙った遺伝子のmRNAまたはたんぱく質の量が減少しているかを調査しました。その結果、ANX 遺伝子ではmRNAの量は減少していませんでしたが、CalS5 遺伝子とROP1 遺伝子ではmRNAの量が減少していました。また、人工的にsGFP 遺伝子を発現させた花粉管でS化オリゴを用いてsGFP遺伝子の働きを抑えた場合には、sGFP遺伝子のmRNAではなく、たんぱく質が減少していることが確認されました。このことから、S化オリゴはmRNAまたはたんぱく質の発現を抑える効果があり、その効果は遺伝子ごとに異なることが示されました。
<今後の展開>
開発したS化オリゴによる阻害方法を用いることで、遺伝子組み換え技術に頼らずにシロイヌナズナの花粉管の発芽、および伸長に必要な遺伝子の働きを狙って抑制する方法が確立されました。「東山ライブホロニクスプロジェクト」では、植物の生殖過程において、花粉管が伸びていく際のめしべとの物質のやり取りなど、植物の受精に関する研究を展開しています。今回開発した方法により、受精における花粉管の役割の研究が、より一層加速されることが期待されます。
また、培地上に散布した花粉管だけでなく、めしべの中を正常に進んできた花粉管でも、切り口から出た後にS化オリゴに出会うことで、その働きが阻害されることも分かりました(図7)。この発見は世界初であり、今後、花粉管の発芽・伸長に必要な遺伝子だけでなく、花粉管とめしべとの物質のやり取りに必要な遺伝子を阻害するなど、受精過程における花粉管の細胞とめしべの細胞の間のコミュニケーションの研究が進むことが期待されます。
被子植物の花粉の多くは、めしべの上で吸水して発芽することから、このS化オリゴによる遺伝子発現の制御方法は、シロイヌナズナだけでなく、他の植物の花粉管にも応用することができます。つまり、これまで花粉管の伸長に問題があり交雑ができなかった植物同士でも、花粉管の伸長を妨げる遺伝子の働きを抑えて受精を成功させるなど、遺伝子組み換えを用いることなく新しい農作物を作ることも可能です。さらに、基礎研究においても、遺伝子組み換え体を作る必要がないことから、研究にかかる所要時間の大幅な削減が期待されます。また、遺伝子組み換え実験を行う設備がない場所での実験も可能になります。
<参考図>
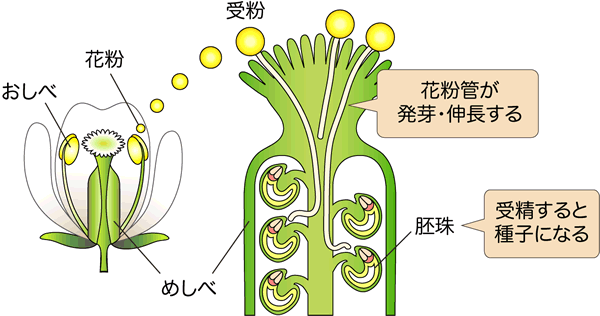
図1 シロイヌナズナの花とめしべ内部の模式図
左:シロイヌナズナの花の模式図、右:シロイヌナズナのめしべ内部の模式図
めしべ上部におしべから花粉が付着(受粉)した後、花粉管が発芽し、めしべ内部へ伸長していきます。この花粉管がめしべ内の胚珠へ到達し、受精が成功することで種子ができます。
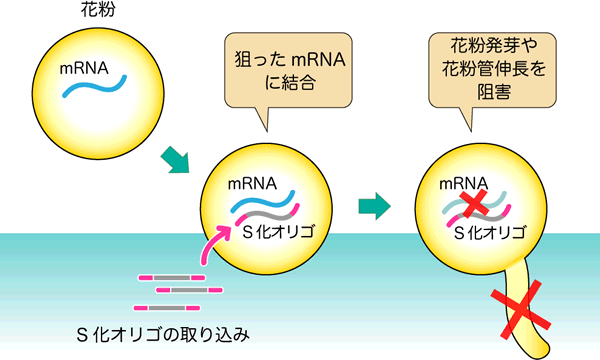
図2 S化オリゴを用いた花粉内の遺伝子発現阻害の原理
シロイヌナズナの花粉は乾燥しているため、花粉を培地に散布すると吸水して花粉管を発芽します。その際、添加したS化オリゴも水分とともに花粉の中に取り込まれます。取り込まれたS化オリゴは狙った遺伝子のmRNAに結合し、その働きを抑えることで、花粉発芽や花粉管伸長が阻害されると考えられます。
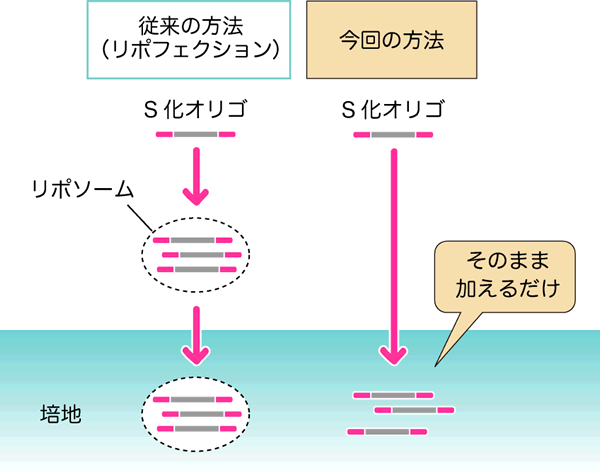
図3 従来の方法と今回の方法の比較
花粉へのS化オリゴの取り込みには、従来はリポフェクションを行ったS化オリゴを培地に加える方法が用いられていました。一方、今回の方法では、S化オリゴをそのまま培地に加えるだけで効果が得られました。
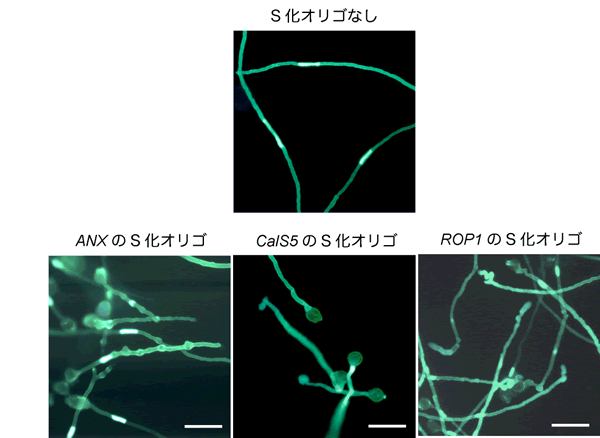
図4 S化オリゴによる花粉管の形態変化
S化オリゴを含まない培地、およびそれぞれの遺伝子に対するS化オリゴを加えた培地で伸長した花粉管の写真です。S化オリゴを含まない培地では、花粉管がまっすぐ伸長しているのに対し、ANX 遺伝子のS化オリゴを加えた培地では花粉管にこぶや枝分かれが観察されました。また、CalS5 遺伝子のS化オリゴでは短い花粉管が、ROP1 遺伝子のS化オリゴでは蛇行し先端が波打っている花粉管が観察されました。S化オリゴが標的とする遺伝子によって、花粉管の形態がそれぞれ異なることが分かります。スケールバーは0.1mmです。
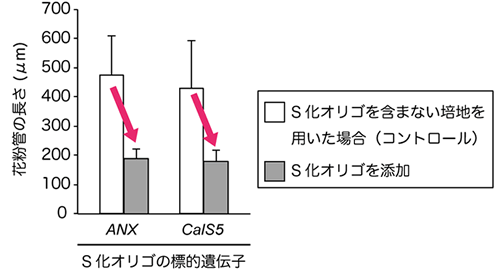
図5 花粉管の長さに影響する遺伝子に対するS化オリゴの阻害効果
白はS化オリゴを含まない培地を用いた場合の花粉管の長さ(コントロール)、灰色はANX 遺伝子、およびCalS5 遺伝子の各S化オリゴを含む培地を用いた場合の花粉管の長さを示します。コントロールに比べて、S化オリゴを加えた場合は、花粉管の長さが顕著に短くなっていることが分かります。
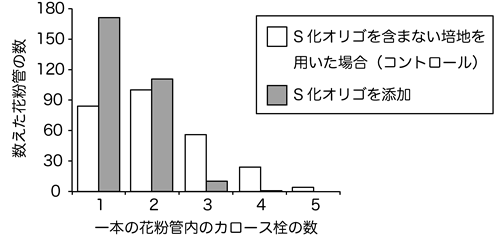
図6 CalS5 遺伝子に対するS化オリゴを用いた際のカロース栓の数
白で示したS化オリゴを含まない培地用いた場合(コントロール)に対し、灰色で示したCalS5 遺伝子のS化オリゴを用いた場合では、花粉管内のカロース栓の数が減少していることが分かります。このことから、CalS5 遺伝子の働きを抑えると、カロースの合成が抑えられ、カロース栓の数が減少すると考えられます。

図7 めしべの切り口から出てきた花粉管に対するS化オリゴの効果
左はめしべの切り口から出てきた花粉管を、S化オリゴを含まない培地で培養したものです。花粉管がまっすぐ伸びているのが分かります。一方、右はANX 遺伝子に対するS化オリゴを加えた培地上で、めしべの切り口から出てきた花粉管を培養したものです。左の写真に比べて、右の写真では花粉管の先端が膨らみ、先端が破裂してしまっているものも観察されました。スケールバーは0.1mmです。
<用語解説>
- 注1)花粉管
- 被子植物では、花粉がめしべに受粉した後、花粉内の精細胞をめしべの生殖細胞(胚珠)へと届けるために、花粉から長い管状の細胞が伸びていきます。これを花粉管と呼びます。花粉管がめしべの深部にある胚珠まで伸びることで受精が起き、種子が形成されます。種子ができるためには、花粉管が正常に発芽・伸長することが必要です。
- 注2)遺伝子組み換え
- 遺伝子操作を行い、新たな遺伝子を導入し発現させたり、もともとその生物が持っている遺伝子の発現を促進・抑制したりすることを遺伝子組み換えと呼びます。また、遺伝子組み換えにより、新たな形質が付与された植物のことを遺伝子組み換え植物と呼びます。研究目的で遺伝子組み換え植物を栽培する場合は、所定の手続きや、定められた栽培環境が必要となります。
- 注3)ホスホロチオエート化
- ホスホロチオエート化はS化、硫黄化ともいう。通常、人工合成したオリゴDNAでは、塩基間のリン酸基がホスホジエステル結合されているのに対し、ホスホロチオエート結合されているものをS化オリゴDNAと呼びます。
- 注4)アンチセンスオリゴDNA
- ゲノム上にある遺伝子からは、mRNAが転写され、mRNAからはたんぱく質が翻訳されることで、遺伝子の情報が細胞へと伝えられます。標的とする遺伝子のmRNAに対し、結合する配列(相補的配列)を持つように人工合成されたDNAオリゴはアンチセンスオリゴDNA(アンチセンスオリゴ)と呼ばれます。
- 注5)交雑育種
- 人工的に植物や動物を掛け合わせて、新しい雑種を作ることを交雑育種と呼びます。2つの種が持つ別々の長所を1つの個体に合わせることができるため、農業分野ではこれまでも盛んに行われてきました。例として、カブとキャベツの掛け合わせ(交雑)からはナタネが作り出されています。一方、イネの品種間などの交雑では、ある特定の遺伝子が妨害をするため雑種を作ることができないなど、交雑を妨げる遺伝子があることも知られています。
- 注6)相補的な配列
- 通常、オリゴDNAはA、T、G、Cの4種類の塩基から構成されているのに対し、mRNAはA、U、G、Cの4種類の塩基から構成されています。オリゴDNAとmRNAが結合する場合、AとU、GとCが結合するという決まりがあります。この性質を利用し、狙った遺伝子のmRNAに完全に結合するような配列を持っている、つまり相補的な配列を持つオリゴDNAはアンチセンスオリゴDNAと呼ばれ、さまざまな実験に利用されています。
- 注7)μM(マイクロモーラー)
- 濃度を表す方式の1つで、モル濃度と呼ばれます。1リットルの溶液中に何マイクロモル(マイクロは100万分の1)の物質が溶けているかを表します。
- 注8)カロース、カロース栓
- カロースは植物が合成する多糖類の一種です。カロース栓は、花粉管内に観察される栓のような構造物で、花粉管の細胞の内側にカロースと呼ばれる物質が蓄積することで形成されます。カロース栓は花粉管が伸長するに伴って定期的に形成され、花粉管の伸長など、正常な受精を行うのに重要な役割を果たしていると考えられています。
<論文タイトル>
“Antisense gene inhibition by phosphorothioate antisense oligonucleotide in Arabidopsis pollen tubes”
(シロイヌナズナの花粉管におけるホスホロチオエート化アンチセンスオリゴを用いた遺伝子発現阻害)
doi: 10.1111/tpj.12461
<お問い合わせ先>
<研究に関すること>
水多 陽子(ミズタ ヨウコ)
名古屋大学 大学院理学研究科 ERATO東山ライブホロニクスプロジェクト 研究員
〒464-8602 愛知県名古屋市千種区不老町
Tel:052-747-6401 Fax:052-789-2497
E-mail:
<JSTの事業に関すること>
坂本 祥純(サカモト ヨシズミ)
科学技術振興機構 研究プロジェクト推進部
〒102-0076 東京都千代田区五番町7 K’s五番町
Tel:03-3512-3528 Fax:03-3222-2068
E-mail:
<報道担当>
科学技術振興機構 広報課
〒102-8666 東京都千代田区四番町5番地3
Tel:03-5214-8404 Fax:03-5214-8432
名古屋大学 広報室
Tel:052-789-2016