ポイント
- 植物の受精では多精拒否の仕組みがあるが、これまでそのメカニズムは謎であった。
- 2つの生殖細胞(卵細胞と中央細胞)が独立して花粉管誘引停止を制御することを発見。
- 別々の花粉と受精する「ヘテロ受精」に成功。新しい雑種を作る技術の応用に道。
JST 課題達成型基礎研究の一環として、名古屋大学 WPI トランスフォーマティブ生命分子研究所の丸山 大輔 研究員、JST 戦略的創造研究推進事業 ERATO型研究「東山ライブホロニクスプロジェクト」の東山 哲也 研究総括(名古屋大学 WPI トランスフォーマティブ研究所 教授)らは、植物が花粉管注1)誘引を停止する仕組みを発見しました。
被子植物では、雄の組織である花粉管に存在する2つの精細胞が、雌の組織である胚珠内の胚のう注2)にある卵細胞と中央細胞それぞれと重複受精注3)します。その結果、植物体となる胚と、栄養を蓄える胚乳を持つ種子が作られます。通常、1つの胚珠は1本の花粉管のみを誘引・受容し、受精後はそのほかの花粉管は誘引しないことが知られています。これは、植物の多精拒否の仕組みの1つと考えられていますが、受精により花粉管誘引が停止する詳しいメカニズムは分かっていませんでした。
今回丸山研究員らは、重複受精できない(通常1本の花粉管につき2つある精細胞が1つしか作られない)シロイヌナズナの変異体を利用して、重複受精と花粉管誘引停止の関係を明らかにしました。変異体では花粉管の誘引を完全には停止できず、約30%の胚のうで複数の花粉管を誘引することが分かりました。この結果から、卵細胞あるいは中央細胞の受精はともに花粉管の誘引停止に関係しており、両方の受精がそろう(重複受精が成功する)ことにより誘引が完全に停止することが示されました。さらに、卵細胞と中央細胞では異なる制御メカニズムを持つと考えられ、この性質を応用することで、ある個体の花粉で卵細胞を受精させたのち、別の個体の花粉で中央細胞を受精させる「ヘテロ受精注4)」を人為的に引き起こすことに成功しました。
ヘテロ受精とは胚と胚乳が異なる遺伝子型を持つようになる受精の様式を指し、トウモロコシで80年以上前に報告されています。しかし、これまでヘテロ受精がなぜ起こるのか明らかではありませんでした。本研究で、ヘテロ受精の背景に花粉管の誘引停止の仕組みが深く関わっていることが明らかとなっただけでなく、任意にヘテロ受精の種子を作り出すことが可能であることが示されました。今後、胚と胚乳の両者に異なる遺伝子を持たせ、両者を最大限に成長させる技術や、種間交雑の際の両者の不和を解消して新しい有用な植物の作出技術などにつながると期待されます。
本研究成果は、2013年5月13日(米国東部時間)に米科学誌「Developmental Cell」オンライン速報版で公開されます。
本成果は、以下の事業・研究プロジェクトによって得られました。
戦略的創造研究推進事業 ERATO型研究
| 研究プロジェクト |
「東山ライブホロニクスプロジェクト」
|
| 研究総括 |
東山 哲也(名古屋大学 WPI トランスフォーマティブ生命分子研究所 教授) |
| 研究期間 |
平成22年~平成27年 |
JSTはこのプロジェクトで、ライブセル解析による多細胞生物のホロニックコミュニケーションの全容解明に挑みます。多細胞生物の細胞は、種々のシグナル分子を介して、ほかの細胞と情報のやり取りをすることで自身の振る舞いを決定し、生物としてのバランスを取っています。こうした個と全体を調和する細胞間コミュニケーションを、ホロニックコミュニケーションと呼びます。そのメカニズムの解明および解析技術の確立は、広く生物学や医療の分野での貢献が期待できます。本研究では、そのための多くのライブセル解析技術の創出を目指します。
<研究の背景と経緯>
被子植物の最もユニークな特徴の1つが、卵細胞と中央細胞という、2つの生殖細胞を独立に受精させて、胚(種子の中で植物体になる部分)と胚乳(種子の中の栄養器官)を作ることです(図1A)。これを重複受精といいます。植物の卵細胞と中央細胞を、異なる遺伝子を持つ精細胞と受精させることができれば、胚と胚乳に異なる遺伝情報を持たせるよう制御することが可能です。被子植物の精細胞は鞭毛を欠き、自ら泳ぐことはできません。このため精細胞は、卵細胞の隣に2つある助細胞により誘引され花粉から伸びる「花粉管」により運ばれます。しかし、1本の花粉管によって同じ遺伝子組成の2つの精細胞が運ばれ、それらが卵細胞、中央細胞と受精するため、胚と胚乳に異なる遺伝情報を持たせるような制御は難しいと考えられてきました。また、受精が完了すると、残りの花粉管が誘引されることはありません。一方で、トウモロコシでは、まれに胚と胚乳が異なる遺伝子を持つ精細胞と受精する「ヘテロ受精」が起こることが、1929年にアメリカのスプラグによりScience誌に報告されています。これまで、どのようにヘテロ受精が起こるのか、その仕組みは解明されていませんでした。このため、胚と胚乳を異なる遺伝子組成にして両者を制御することは原理的には可能であるものの、人為的な制御は難しいと考えられてきました。
<研究の内容>
丸山研究員らは、重複受精における2つの雌しべ側の生殖細胞(卵細胞と中央細胞)の受精と、花粉管の誘引停止の関係を明らかにするために、アブラナ科植物であるシロイヌナズナを用いて調べました。被子植物の重複受精は、雌しべの奥深くでほぼ同時に起きるために、卵細胞と中央細胞の受精を区別して解析することは困難でした。そこで、通常2つある精細胞が1つしか作られない突然変異のあるシロイヌナズナcdka;1 変異体を利用しました。この変異体の花粉を授粉させると、卵細胞と中央細胞のどちらか一方のみと受精が起こるため、胚または胚乳のどちらかのみが発達した種子が作られます。これらの種子を調べたところ、どちらのタイプでも花粉管の誘引は完全には停止せず、約30%の割合で複数の花粉管を誘引していることが分かりました。こうした解析から、以下のような関係性があると考えられました(図2)。
- ① 卵細胞と中央細胞は、それぞれ受精することで2本目の花粉管の誘引を止めるように作用するが、どちらか一方の受精だけでは不十分で、花粉管の誘引を完全に止めることができない。
- ② 卵細胞と中央細胞が両方とも受精すると、協調的に作用し2本目の花粉管誘引を停止できる。
また、卵細胞と中央細胞での花粉管誘引の制御は独立して起こり、中央細胞では胚乳形成を制御する遺伝子としてよく知られていたMEDEA 遺伝子が、花粉管の誘引停止に必要であることも分かりました。
これらの結果から、今度は卵細胞または中央細胞の片方だけが受精に成功した雌しべ側の生殖細胞について、次に誘引された花粉管で重複受精が完了する、すなわちヘテロ受精が起こるのではないかと考え、解析を進めました。実験には、2つの精細胞のうち片方だけが受精する傾向を示す突然変異体のシロイヌナズナkokopelli 変異体を用いました。kokopelli 変異体に、細胞核が顕微鏡下で赤色に光るようにさらに改変を加えた変異体を作り、その花粉を正常なシロイヌナズナに授粉しました。また、その3時間後に、今度は細胞核が顕微鏡下で緑色に光るように改変した変異体の花粉を授粉しました。これらの2種類の変異体を利用し時間差をつけて花粉を授粉させることにより、初めのkokopelli 変異体の授粉で卵細胞または中央細胞のいずれかと受精し、次の授粉で残った方の受精が起きます。このことにより、胚と胚乳の細胞核の色が異なる種子が観察できると考えられました。実験の結果、胚と胚乳の細胞核の色が異なる種子を得ることに成功しました(図1B、C)。シロイヌナズナでヘテロ受精が示されたのは初めてのことです。また、ヘテロ受精を人為的に引き起こすことができることも明らかとなりました。これまで謎だったヘテロ受精には、卵細胞と中央細胞のそれぞれの受精による協調的な花粉管誘引停止の仕組みが背景にあることが分かりました。
<今後の展開>
今回の研究成果をもとに、卵細胞と中央細胞のそれぞれがどのように協調的に花粉管誘引を停止するのか、その仕組みの解明が期待されます。今回の成果は、ドイツのグロスハート博士らの論文と連報で誌面を飾ります。グロスハート博士らは、花粉管の誘引停止には、誘引をつかさどる助細胞という細胞にエチレンが作用して細胞死を引き起こすことが必要であることを示しました。卵細胞や中央細胞の受精がどのようにエチレンの生成に関わるのか、今後の研究の展開が期待されます。
また、ヘテロ受精を人為的に起こすことができるようになったことは、基礎研究と応用研究の両面での展開が期待されます。胚と胚乳は種子の中で協調して形成が進行します。その際の胚と胚乳の間での生成物のやり取り(コミュニケーション)が両者の発達に対して極めて重要であることが知られていますが、これまでは、それぞれを異なる遺伝子組成にすることが不可能であったために、胚と胚乳で生成される物質の区別をつけることが難しく、どのようなコミュニケーションが行われているのか、具体的には明らかになっていません。今回の研究成果によりヘテロ受精を人為的に制御できるようになったことで、胚と胚乳の相互作用による種子形成の仕組みの解明が期待されます。
応用面では、卵細胞と中央細胞の受精を独立に制御することで、さまざまな展開が期待されます。例えば、胚と胚乳を異なる発生プログラムで成長するため、それぞれに最適化した遺伝子を持たせることで、良く発達した、貧栄養の土地でも育ちやすい種子を得ることも可能になると期待されます。また、育種において異なる植物種を交雑させ新しい植物種を得る際に、胚と胚乳のコミュニケーションがうまくいかないために種子形成ができず、雑種(新しい植物種)が得られないことがあります。こうした不和を取り除いたり、あるいは雑種の胚の成長をサポートするような胚乳を作り出すことで、新しい有用な植物の作出につながる可能性も考えられます。このように今回の研究成果でヘテロ受精の謎が解明され、人為的な制御が可能になったことで、植物の生殖制御技術の新しい扉が開いたといえます。
<参考図>
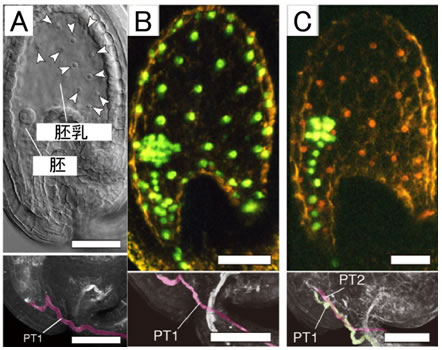
図1 重複受精による胚と胚乳の形成と、ヘテロ受精の人為的な制御
- (A)重複受精により、種子の内部に胚と胚乳が形成される。
- (B)重複受精は同じ遺伝子組成を持つ1対の精細胞により達成される。そのため、緑色に光るタンパク質を発現するような精細胞で受精が起こると、胚も胚乳も緑色に光る。
- (C)kokopelli 変異体を用いて人為的にヘテロ受精を起こすことを試みた結果、緑色に光るタンパク質を発現する精細胞と、赤色に光るタンパク質を発現する精細胞で受精が起こり、胚と胚乳が異なる色で光る種子が得られた。
それぞれの下段では、上段と同じ組織における花粉管(PT)を可視化している。(C)では花粉管が2本到達していることが確認できる。
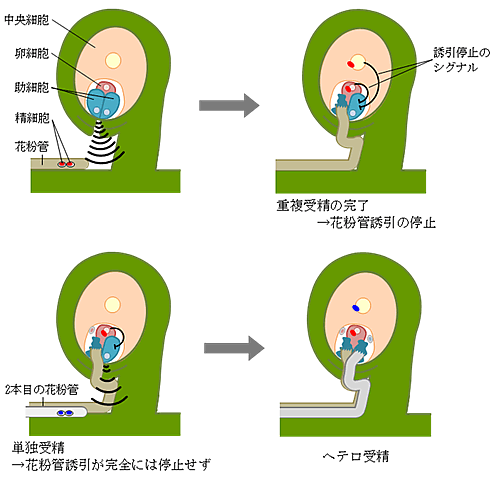
図2 卵細胞と中央細胞による花粉管の誘引停止の模式図
卵細胞と中央細胞の受精がそれぞれ独立に、花粉管を誘引する助細胞に作用し、花粉管の誘引が完全に停止する(上段)。片方の受精(単独受精)でも、誘引の低下が見られるものの、花粉管誘引は完全には停止しない(下段左)。このため、2本目の花粉管が誘引され、ヘテロ受精が起こる(下段右)。
<用語解説>
- 注1) 花粉管
- 花粉が雌しべに受粉した後、精細胞を雌しべ深部にある生殖細胞へと届けるために花粉から伸びる長い管状の細胞。先端の内部に2個の精細胞を持つ。花粉管が雌しべ側の生殖細胞に到達することで受精が起き、種子が形成される。
- 注2) 胚のう
- 多くの場合、1個の卵細胞、1個の中央細胞、2個の助細胞と3個の反足細胞からなる、受精後に種子を形作る中心的組織。
- 注3) 重複受精
- 卵細胞と中央細胞の2つの細胞が受精をする被子植物に独自の受精様式。
- 注4) ヘテロ受精
- 胚と胚乳が異なる遺伝子型を持つようになる特殊な型の受精現象。
<論文名>
“Independent control by each female gamete prevents the attraction of multiple pollen tubes”
(各雌性配偶子による独立の制御が複数の花粉管誘引を阻止している)
doi: 10.1016/j.devcel.2013.03.013
<お問い合わせ先>
<研究に関すること>
東山 哲也(ヒガシヤマ テツヤ)
名古屋大学 ERATO 東山ライブホロニクスプロジェクト 研究総括
〒464-8602 愛知県名古屋市千種区不老町
Tel:052-747-6404 Fax:052-789-2497
E-mail:
<JSTの事業に関すること>
坂本 祥純(サカモト ヨシズミ)
科学技術振興機構 研究プロジェクト推進部
〒102-0076 東京都千代田区五番町7 K’s五番町
Tel:03-3512-3528 Fax:03-3222-2068
E-mail: