千葉大学 大学院理学研究科の村田 武士 特任准教授(JST さきがけ研究者、理化学研究所 客員研究員)らは、たんぱく質ナノモーター注1)であるV型ATPaseの回転分子モーター部分の詳細構造を世界で初めて解明しました。これにより、ATPのエネルギーが回転運動に変換される仕組みの大枠が原子レベルで明らかになりました。骨粗鬆症やがんなどの疾病に関与するV型ATPaseを阻害する方法の予測が可能となり、立体構造に基づいた治療薬の創製に繋がるものと期待されます。
本研究は文部科学省ターゲットタンパク研究プログラム、文部科学省 科学技術振興調整費、JST 戦略的創造研究推進事業 さきがけなどの支援を受け、東京理科大学 大学院基礎工学研究科の山登 一郎 教授、京都大学 大学院医学研究科の岩田 想 教授、理化学研究所 生命分子システム基盤研究領域の横山 茂之 領域長らとの共同研究として行われました。本研究成果は、2013年1月13日(英国時間)の英国科学雑誌「Nature」のオンライン速報版で公開されます。
<研究の背景と経緯>
V型ATPaseは、細菌からヒトまで多くの生体膜中に存在し、ATPのエネルギーを使って水素イオンを運ぶことで膜内外のpHを調整しています。V型ATPaseは骨の形成に関わる破骨細胞やがん細胞の細胞膜にも存在しており、骨粗鬆症やがん細胞の増殖・転移に関与していることが分かっています。そのため、V型ATPaseの分子メカニズムを知ることは、これら疾病の理解に繋がりますし、V型ATPaseの阻害剤は治療薬として期待されています。V型ATPaseは、親水性のV1部分と膜内在性のVo部分から構成されています(図1)。触媒頭部(A3B3複合体)でATPが加水分解され、そのエネルギーを使って軸部分が回転し、これに伴ってVo部分で水素イオンが輸送されると考えられていますが、詳細構造が不明であったため詳しい仕組みは未解明でした。当研究グループは、細菌(腸球菌)にもヒトV型ATPaseに良く似た酵素が存在することを発見し、その生化学的・構造生物学的研究を進めてきました。最近、ヒトと腸球菌のV型ATPaseの回転軸(DF複合体)の詳細構造をそれぞれ明らかにしました。得られた構造は互いにそっくりで、V型ATPaseの動作原理はヒトと腸球菌で酷似していることを明らかにしています。本研究では、V型ATPaseがATPの化学エネルギーを物理的な回転エネルギーに変換する仕組みを明らかにすることを目的に、腸球菌V型ATPaseの触媒部分及びV1複合体のX線結晶構造解析注2)を試みました。
<研究の内容>
当初は大腸菌を用いてAとBサブユニットをそれぞれ精製後、A3B3複合体を再構成していましたが、大量調製が難しく、良質の結晶を得ることができませんでした。そこで、大腸菌無細胞たんぱく質合成系を用いてAとBサブユニットの共発現精製系を検討し、A3B3複合体の大量精製に成功しました。この高純度な標品を用いることにより、ATPなどのヌクレオチドは入れない条件で良質の結晶を得ることができ、そのX線結晶構造(分解能2.8Å)を明らかにしました(図2a)。得られた複合体リング構造は、同じ3つのAサブユニット、Bサブユニットから構成されているのにも関わらず完全に非対称であり、それぞれのサブユニットはすべて異なる立体構造を形成していました(図2b)。
次に、ATPに似た構造をもつAMP-PNP注3)存在下で、A3B3複合体のX線結晶構造(分解能3.4Å)を明らかにしました。A-B境界面に存在する計3ヵ所のATP結合部位のうち、2ヵ所にAMP-PNPが結合し、これにより一部のA3B3複合体の立体構造が変化していました(図2c、d)。得られた2つのA3B3複合体の構造を比較することにより、A3B3複合体は「ATPが結合できないフォーム:Empty」、「ATPを結合することができるフォーム:Bindable」、「ATPを結合しているフォーム:Bound」の3つの異なるA-B複合体から構成されていることが明らかになりました(図2b)。ATP存在下では、BindableフォームにATPが結合し、2つのBoundフォームができます(図2d)。元々あったBoundフォームのATPが分解されると、A3B3複合体は初めの構造に戻るように変化すると考えられます。つまりA3B3複合体の構造を120°回転させた構造に変化することになります。以上の考察から、なぜ分子モーターがATPのエネルギーを使って一方向に回転するかの仕組みを立体構造から理解することができました。
次に、A3B3複合体とDF複合体からV1-ATPase(A3B3DF複合体)の再構成条件をSPR(表面プラズモン共鳴)法注4)を用いて検討しました。酸性pHやマグネシウム存在下でV1-ATPaseを安定に精製することができ、この標品を用いることでV1-ATPaseの高分解能X線結晶構造(2.2Å)を得ることに成功しました(図3a)。上記のA3B3複合体と構造を比較することにより、DF複合体結合によるA3B3複合体の構造変化を理解することが可能となります。その結果、DF複合体が結合することにより、「ATPを結合することができるフォーム:Bindable」はATPが結合していないのに「ATPを結合しているフォーム:Bound」に変化し、「ATPを結合しているフォーム:Bound」はA3B3複合体には存在しなかった「よりコンパクトなフォーム:Tight」に変化していました(図3b)。さらに、AMP-PNPが2つ結合したV1-ATPaseのX線結晶構造(2.7Å)を明らかにしました。これらの構造を比較することにより、結合したATPは「よりコンパクトなフォーム:Tight」で加水分解が誘導されることが示唆されました。つまり、DF複合体は単にA3B3複合体の構造変化に応じて回転しているのではなく、A3B3複合体と結合して構造を変化させて、ATPの分解場所を決定していることが示唆されました。以上の発見を基に全く新規なV1-ATPaseの回転メカニズムモデルを提案することができました。
<今後の展開>
本研究により、A3B3複合体の役割、DF複合体の役割、なぜ一方向に回転するかの理由、ATPがどこで分解されるかなど、多くの情報を理解することができるようになりました。これにより、その他のたんぱく質ナノモーターを含む生体エネルギー変換機構の一般原理の解明に繋がることが期待されます。また、V1-ATPaseの詳細な分子メカニズムが解明されることで、関連する骨粗鬆症やがん転移などの疾病原因の理解やその治療薬の開発についても進展することが期待されます。
<参考図>
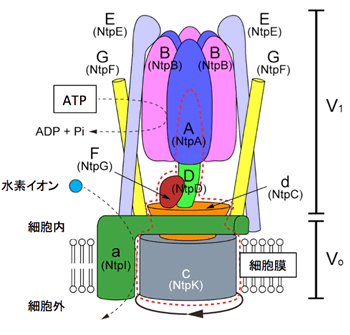
図1 V型ATPaseの構造モデル
V型ATPaseは9-13種類のたんぱく質からなる超分子複合体で、水溶性たんぱく質部分(V1部分)と膜たんぱく質部分(Vo部分)からなる。括弧内の名称は腸球菌V-ATPaseサブユニットの旧名。触媒頭部(A3B3)でATPを加水分解し、回転軸(DFd)とローターリング(c)を回転させ、水素イオンを細胞外(またはオルガネラ内)へ輸送する。
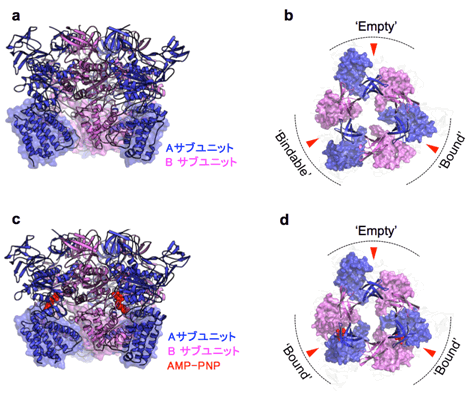
図2 触媒部分(A3B3複合体)のX線結晶構造
- a)横から見たA3B3複合体の結晶構造。
- b)上から見たA3B3複合体の結晶構造。見やすくするためにN末βバレルドメインとC末ドメインのみを表示している。3ヵ所あるATP結合部位を赤い矢印で示した。
- c)、d)AMP-PNPが結合したA3B3複合体の結晶構造。a)、b)と同様に表示。
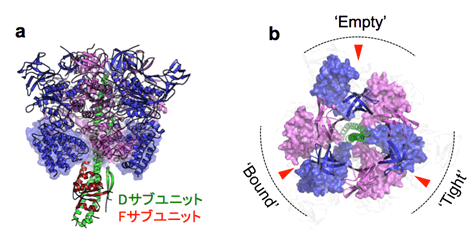
図3 V1-ATPase(A3B3DF複合体)のX線結晶構造
- a)横から見たV1-ATPaseの結晶構造。
- b)上から見たV1-ATPaseの結晶構造。見やすくするためにN末βバレルドメインとC末ドメイン、Dサブユニットのみを表示している。3ヵ所あるATP結合部位を赤い矢印で示した。
<用語解説>
- 注1) たんぱく質ナノモーター
- 生物の細胞内でATPの加水分解によって得られたエネルギーを、機械的な運動に変換するナノメートル(10-9m)程度のたんぱく質複合体の総称。
- 注2) X線結晶構造解析
- 解析対象のたんぱく質を結晶化し、X線照射によって得られる回折データから、たんぱく質の原子レベルでの立体構造を決定する手法。
- 注3) AMP-PNP
- ATPの非分解性の類似体(アナログ)。β-とγ-リン酸間が窒素原子に置き換わっているため、加水分解反応が阻害される。
- 注4) SPR(表面プラズモン共鳴)法
- センサー表面にたんぱく質を固定し、他の分子との相互作用を標識なしに高感度かつリアルタイムに測定できる手法。
<論文名および著者名>
雑誌名:Nature
論文名:“Rotation mechanism of Enterococcus hirae V1-ATPase based on asymmetric crystal structures”
著者:Satoshi Arai, Shinya Saijo, Kano Suzuki, Kenji Mizutani, Yoshimi Kakinuma, Yoshiko Ishizuka-Katsura, Noboru, Ohsawa, Takaho Terada, Mikako Shirouzu, Shigeyuki Yokoyama, So Iwata, Ichiro Yamato, and Takeshi Murata
doi: 10.1038/nature11778
<お問い合わせ先>
<研究に関すること>
村田 武士(ムラタ タケシ)
千葉大学 大学院理学研究科 特任准教授
Tel:043-290-2794
E-mail:
<JSTの事業に関すること>
石正 茂(イシマサ シゲル)
科学技術振興機構 戦略研究推進部 ライフイノベーショングループ
Tel:03-3512-3524
E-mail: