ポイント
- 敗血症は日本で年間38万人が発症し、死亡率が高い
- 免疫細胞の1つ、肥満細胞の敗血症を抑制する働きをたんぱく質「MAIR-Ⅰ」がコントロール
- MAIR-Ⅰの働きを抑えたマウスでは敗血症の生存率が40%に増加。革新的な治療薬へ期待
JST 課題達成型基礎研究の一環として、筑波大学 医学医療系の渋谷 彰 教授と小田 ちぐさ 助教らは、敗血症の発症に関与する免疫受容体注1)を発見し、新しい敗血症の予防と治療につながる手法の開発に成功しました。
敗血症は、細菌による感染を発端として、細菌が産生する毒素が全身に広がり、多臓器不全、血圧低下、ショックなどの症状を引き起こす重篤な全身疾患です。腹膜炎や肺炎、術後感染などからの敗血症の発症数は世界的に増えつつあり、その死亡率は非常に高く、新たな治療法が望まれる重要な疾患です。日本では年間38万人以上が発症すると推定されています。
細菌に感染すると、免疫細胞の一種である肥満細胞注2)が感染局所でそれをいち早く感知し、白血球の一種の顆粒球注3)を感染局所に動員して細菌を直接死滅させることで、敗血症の発症を抑制することが知られていました。しかし、肥満細胞の働きがどのようにコントロールされているか、また、顆粒球が存在するにも関わらず敗血症が発症する理由については、十分には明らかにされていませんでした。
今回本研究グループは、肥満細胞の細胞表面膜たんぱく質である免疫受容体「MAIR-Ⅰ」(別名CD300a)が、肥満細胞の顆粒球を動員する働きを抑えることにより、敗血症の発症を促進することを明らかにしました。同時に、MAIR-Ⅰの働きを抑えた遺伝子欠損マウスでは敗血症の発症が抑制され、生存率が著明に増加(0%→40%)することを突き止めました。また、抗体などのたんぱく質でMAIR-Ⅰの働きを抑えた場合にも、生存率の増加が見られました。これらの結果は、革新的な敗血症治療薬開発につながるものとして期待されます。
本研究は、京都大学 長田 重一 教授、大阪大学 菊谷 仁 教授らの協力を得て行われ、2012年7月23日(米国東部時間)に科学誌「Journal of Experimental Medicine」のオンライン速報版で公開されます。
本成果は、以下の事業・研究領域・研究課題によって得られました。
戦略的創造研究推進事業 チーム型研究(CREST)
| 研究領域 |
「アレルギー疾患・自己免疫疾患などの発症機構と治療技術」
(研究総括:菅村 和夫 宮城県立病院機構 理事長)
|
| 研究課題名 |
「ヒト肥満細胞活性化制御技術の開発によるアレルギー疾患の克服」 |
| 研究代表者 |
渋谷 彰(筑波大学 医学医療系 教授) |
| 研究期間 |
平成22年10月~平成28年3月 |
JSTはこの領域で、アレルギー疾患や自己免疫疾患を中心とするヒトの免疫疾患を予防・診断・治療することを目的に、免疫システムを適正に機能させる基盤技術の構築を目指しています。
上記研究課題では、ヒト肥満細胞の活性化制御技術を開発することでアレルギーをはじめとする炎症性疾患を予防・治療する方法の開発を目指します。
<研究の背景と経緯>
免疫受容体の働きは、免疫細胞の細胞膜に存在し、外界からの刺激や情報をキャッチして細胞内に伝えることです。病原体や異物が体内に侵入すると、多種多様な免疫細胞が互いに連携してこれらを排除しますが、そのためには特定の刺激や情報をそれに対応した免疫受容体がキャッチして免疫反応を起こすことが必要です。本研究グループは、免疫受容体の研究を通して、免疫反応を解明し、免疫疾患の克服を目指してきました。
その成果の1つとして本研究グループは、肥満細胞などの細胞膜上に発現する新しい免疫受容体を2003年に世界に先駆けて発見し、これをMAIR-I(別名CD300a)と名付けました(参考文献1)。しかし、この免疫受容体の生体内での働きについては、世界中の多くの研究グループによる精力的な研究にも関わらず、不明なままでした。本研究グループは、MAIR-Iがどのような情報をキャッチするかを明らかにするため、MAIR-Iがアポトーシス注4)で死んだ細胞の細胞膜上に発現するリン脂質、フォスファチジルセリン(PS)と結合することを突き止め、2011年に世界に先駆けて報告しました(参考文献2)。その結果、MAIR-Iはアポトーシスによって生じた死細胞の出現を感知し、免疫反応をコントロールする免疫受容体であることが判明しました(図1)。
一方、敗血症とは、細菌などの病原体感染が全身に広がり、細菌由来のエンドトキシン(LPS)などの毒素が原因となって全身性の炎症反応が生じることで多臓器不全、血圧低下、ショックなどの症状が引き起こされる状態です。通常、感染症に対しては、抗生物質などによる治療を行いますが、敗血症に至ると有効な手立てがなく、高い確率で死に至ります。日本における同疾患の発症は年に38万人以上と推計され、米国では毎年およそ100万人の患者が発症し、その20-30%が死亡しています。集中治療が必要な患者の死因としては最多の疾患で、新たな治療法の確立が待たれています。
これまで、肥満細胞が化学物質を放出して敗血症発症の抑制に重要な働きをすることは分かっていましたが(参考文献3)、その仕組みが破綻して敗血症が発症してしまう理由はよく分かっていませんでした。
<研究の内容>
本研究グループは、MAIR-Iが生体内でどのような働きをするかを解明するために、MAIR-Iを産生できない遺伝子欠損マウス(欠損マウス)を作りました。MAIR-Iは死細胞を感知すること、MAIR-Iが発現する肥満細胞は敗血症発症に重要な働きをすること(参考文献3)に着目し、MAIR-Iと敗血症発症との関連を調べました。腹膜炎は腹腔と腹部の臓器を覆っている腹膜に何らかの原因で細菌が感染し、死細胞が多数出現して炎症を起こす疾患で、進行すると敗血症になります。野生型マウスと欠損マウスで腹膜炎による敗血症を誘導し(腹膜炎-敗血症モデル注5))、両者の生存期間を比較しました。その結果、MAIR-Iを産生する野生型マウスでは、腹膜炎発症80時間後の生存率が約10%、100時間後には全て死亡したのに対し、MAIR-Iを産生しない欠損マウスでは腹膜炎発症80時間後の生存率は約60%、100時間後は約40%で、生き残ったマウスはその後回復して死亡することはありませんでした(図2)。この際、欠損マウスの感染場所である腹腔内では、野生型マウスの腹腔内より細菌を直接死滅させる顆粒球が増加しており、実際に細菌の数が減少していました(図3)。
欠損マウスで敗血症発症後の生存率が高かったメカニズムを明らかにするために、野生型マウスと欠損マウスから肥満細胞を取り出し、敗血症を引き起こす原因となる毒素LPS、およびMAIR-Iと結合するPSを細胞表面に持つ死細胞と一緒に培養しました。その結果、欠損マウス由来の肥満細胞は野生型マウス由来の肥満細胞より、顆粒球を動員する働きのあるケモカイン(たんぱく質)を多量に産生していました(図4)。さらに、MAIR-Ⅰの働きを抑えるたんぱく質MFG-E8注6)を加えると、欠損マウス由来と野生型マウス由来の肥満細胞からのケモカインやサイトカインの産生量は同等になりました(図5)。このことから、肥満細胞に存在するMAIR-Ⅰは死細胞のPSと結合し、顆粒球を動員する働きのあるケモカインの産生を抑制していることが分かりました。
そこで、本研究グループは肥満細胞のMAIR-Ⅰと死細胞のPSの結合をブロックできれば、敗血症を抑えることが可能ではないかと考えました。そのために、野生型マウスで、腹膜炎を起こす直前と、腹膜炎発症18時間後にMAIR-Iに対する中和抗体注7)を腹腔内に投与しました。比較実験として投与した対照抗体においても生存率が若干向上したものの、中和抗体を投与したマウスでは対照抗体を投与したマウスに比べて約35%から約80%へと増加しました(図6)。同様にMAIR-Iと死細胞のPSへの結合をブロックするMFG-E8を投与すると、腹膜炎発症80時間後の生存率が約10%から約40%へと約4倍に上昇しました(図7)。
<今後の展開>
敗血症は手術に伴って起こるなど、その発症が予見される場合も多いことから予防法の確立は重要です。本研究成果により、世界中で毎年100万人以上が死亡する敗血症の予防や治療にはMAIR-Ⅰの働きを抑える中和抗体またはMFG-E8が有効な治療薬として期待されます。
また、肥満細胞は抗原抗体反応の結果としてヒスタミンなどの化学物質を放出することで、アレルギー反応を引き起こす原因にもなります。アレルギー反応が起こっている部位では、多くの細胞がアポトーシス死を起こしており、MAIR-Ⅰは過剰なアレルギー反応を抑える上で重要な役割をしている可能性も示唆されます。その点で、MAIR-Ⅰはアレルギーの治療や予防のターゲットにもなることが期待されます。
<参考図>
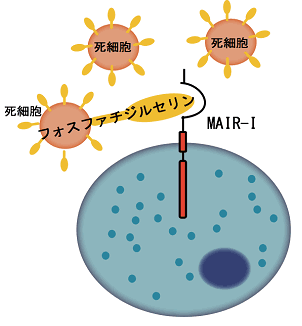
図1 MAIR-Iは死細胞上のフォスファチジルセリンと結合する。
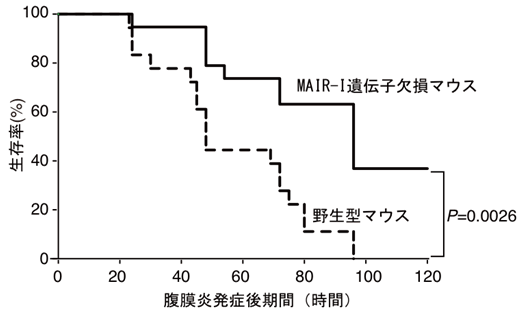
図2 MAIR-I遺伝子欠損マウスでは敗血症による生存率が改善する。
野生型、およびMAIR-I遺伝子欠損マウスに盲腸を結紮穿孔して腹膜炎を誘導し、敗血症を引き起こさせ、生存率を観察するとMAIR-I遺伝子欠損マウスでは生存率が改善した。
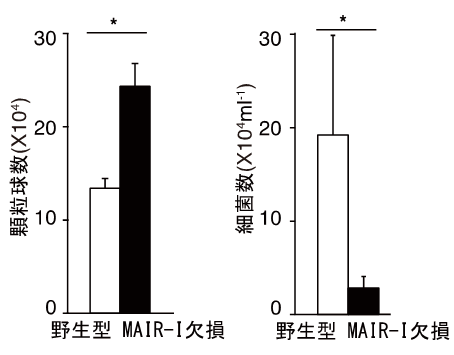
図3 MAIR-I遺伝子欠損マウスでは感染局所での顆粒球の数が増加し、細菌の数が減少する。
腹膜炎をきたしているマウスの腹腔中の顆粒球数、および細菌数を測定した。
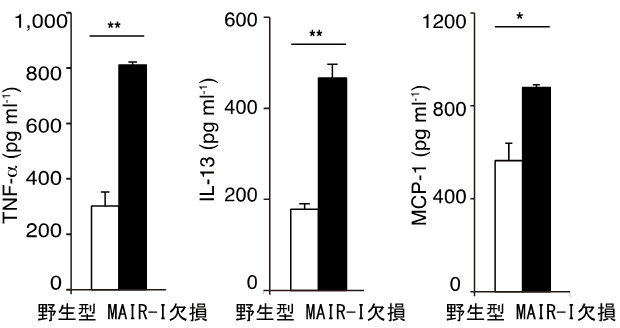
図4 MAIR-I遺伝子欠損マウス由来の肥満細胞からは
多量のケモカインやサイトカインが産生されている。
野生型、およびMAIR-I遺伝子欠損マウスの骨髄から肥満細胞を誘導し、死細胞、およびLPSと共培養し、培養上清中のTNF-α、IL-13、MCP-1を測定した。
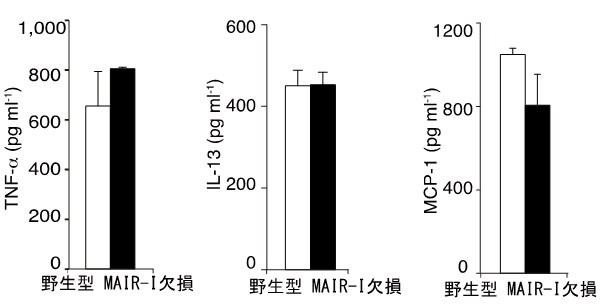
図5 MAIR-Iとフォスファチジルセリンの結合をブロックすると
野生型マウスでもケモカインやサイトカインの産生が増加する。
図4と同様に肥満細胞を誘導し、死細胞とLPSと共培養する際に、MFG-E8を加え、培養上清中のTNF-α、IL-13、MCP-1を測定した。
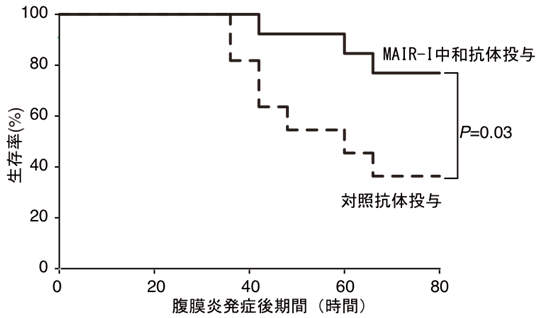
図6 MAIR-Ⅰ中和抗体の投与により敗血症による生存率が改善する。
野生型マウスにMAIR-Ⅰ中和抗体と比較用の対照抗体を投与し、腹膜炎から敗血症を誘導して生存率を観察した。MAIR-Ⅰ中和抗体を投与したマウスでは生存率が顕著に改善した。
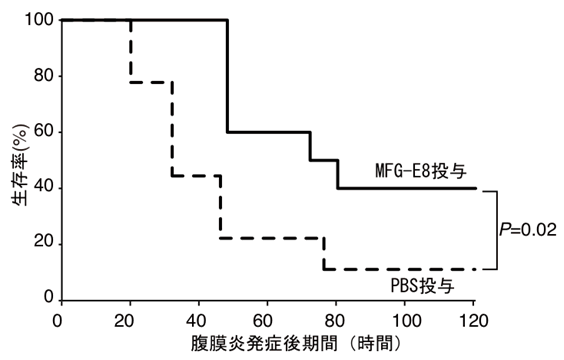
図7 MAIR-Iとフォスファチジルセリンの結合をブロックすると敗血症による生存率が改善する。
野生型マウスにMFG-E8と比較としてPBSを投与し、腹膜炎から敗血症を誘導して生存率を観察した。MFG-E8を投与したマウスでは生存率が改善した。
<参考文献>
1.Yotsumoto, K., Y. Okoshi, K. Shibuya, S. Yamazaki, S. Tahara-Hanaoka, S. Honda, M. Osawa, A. Kuroiwa, Y. Matsuda, D.G. Tenen, A. Iwama, H. Nakauchi, and A. Shibuya. 2003. “Paired activating and inhibitory immunoglobulin-like receptors, MAIR-I and MAIR-II, regulate mast cell and macrophage activation.” J Exp Med 198:223-233.
2.Nakahashi-Oda, C., S. Tahara-Hanaoka, S.I. Honda, K. Shibuya, and A. Shibuya. 2011. “Identification of phosphatidylserine as a ligand for the CD300a immunoreceptor.” Biochem Biophys Res Commun
3.Echtenacher, B., D.N. Mannel, and L. Hultner. 1996. “Critical protective role of mast cells in a model of acute septic peritonitis.” Nature 381:75-77.
<用語解説>
- 注1) 免疫受容体
- 免疫細胞の細胞膜、細胞質または核内にあるたんぱく質で、特異的な物質(リガンド)と結合して免疫反応を開始させる。
- 注2) 肥満細胞
- 免疫細胞の一種でマスト細胞ともいう。全身の粘膜下や結合組織中に存在し、ヒスタミンなどの化学物質を放出することで炎症やアレルギー応答に重要な役割を持つ。
- 注3) 顆粒球
- 白血球の60%を占め、貪食作用や殺菌能力を持ち、細菌からの防御の役割を果たす。好中球、好酸球、好塩基球の3種類がある。
- 注4) アポトーシス
- 細胞の死に方の1つの形態。プログラムされた細胞死ともいわれ、個体にとって余分な細胞や、危険な細胞が個体の持つ制御機構に従って自殺する時の細胞の死に方。生理的にも毎日、全身の多数の細胞がアポトーシスにより死んで新しい細胞によって置き換わっている。細菌などの感染による炎症反応によってもアポトーシスによる細胞死が起こる。
- 注5) 腹膜炎-敗血症モデル
- 虫垂(盲腸)炎から続発する腹膜炎、敗血症マウスモデル。マウスの盲腸を結紮(けっさつ)し、その先端に針で穴をあける。その結果、腹腔へ便が持続的に漏れ、便中の主に大腸菌により腹膜炎をきたし、その炎症が全身に及んで敗血症に至る。
- 注6) MFG-E8
- Milk Fat Globular Protein EGF-8の略。マクロファージが分泌する463個のアミノ酸からなるたんぱく質。アポトーシス細胞の細胞膜に出現するリン脂質であるフォスファチジルセリンに結合し、MAIR-Iとフォスファチジルセリンとの結合を阻止することによってMAIR-Iの働きを抑えることができる。
- 注7) 中和抗体
- 特定の抗原または、抗原の受容体に結合して、抗体抗原反応を起こさせなくする抗体のこと。
<論文タイトル>
“Apoptotic cells suppress mast cell inflammatory responses via the CD300a immunoreceptor.”
(アポトーシス細胞は、CD300a免疫受容体を介して肥満細胞の炎症反応を抑制する)
doi: 10.1084/jem.20120096
<お問い合わせ先>
<研究に関すること>
渋谷 彰(シブヤ アキラ)
筑波大学 医学医療系 教授
〒305-8575 茨城県つくば市天王台1-1-1
Tel:029-853-3281 Fax:029-853-3410
E-mail:
<JSTの事業に関すること>
石井 哲也(イシイ テツヤ)
科学技術振興機構 戦略研究推進部
〒102-0076 東京都千代田区五番町7 K’s五番町
Tel:03-3512-3524 Fax:03-3222-2064
E-mail: