ポイント
- 長期記憶の形成には、神経細胞の情報伝達効率を変化させることが必要。
- 神経細胞で作られるArcたんぱく質が、過剰な神経伝達変化を抑えている可能性。
- 今回のメカニズムは、脳の機能維持や学習・記憶形成に関与。精神疾患などの病態解明へ期待。
JST 課題達成型基礎研究の一環として、東京大学 大学院医学系研究科の尾藤 晴彦 准教授と奥野 浩行 助教らは、マウスやラットを用いた実験により、神経細胞が外部からの刺激に応じて情報伝達効率を調節する新しい分子メカニズムを解明しました。
記憶は、神経細胞のシナプス注1)での情報伝達効率の変化により保存されていますが、その変化は通常、数分から数時間で消失してしまいます(短期記憶)。しかし、強烈な経験や何度も同じ経験をすると、情報伝達効率の変化は数日以上の長期にわたり維持され、長期記憶が形成されます。これは、神経細胞が外部からの刺激に応じてシナプスの性質を長期的に変化させるためであり、こうした仕組みが脳の柔軟性や記憶の形成・保持に必要であると考えられています。これまでの研究により、シナプスの長期変化には、神経細胞の細胞体で新規遺伝子の発現が必要なことが明らかになっています。しかし、神経活動によって発現された遺伝子の産物が細胞のどの部位で、どのような方法で神経細胞の性質を調節し変化させているのかについては、これまでほとんど分かっていませんでした。
本研究グループは、神経活動で発現される遺伝子の産物の1つであるArc(アーク)というたんぱく質に着目しました。Arcたんぱく質はシナプス部位に運ばれ集積しますが、活動性の高いシナプスではなく、逆に活動性の低いシナプスへ運ばれ集積されていることが明らかになりました。さらに、その集積度は神経伝達を担っているグルタミン酸受容体の量と逆相関であることが分かりました。このことから、長期的な記憶が形成されるにあたって、Arcたんぱく質が不要なシナプスの機能を抑える役割を果たしていると考えられます。
ヒトにおいても、このような遺伝子発現による神経機能調節機構は脳の発達や学習・記憶形成などに必須なメカニズムであると考えられます。今回の発見は、精神疾患や学習・記憶障害などの病態解明および治療法の開発につながるものと期待されます。
本研究成果は、2012年5月11日付けの米国科学雑誌「Cell」で公開されます。
本成果は、以下の事業・研究領域・研究課題によって得られました。
戦略的創造研究推進事業 チーム型研究(CREST)
| 研究領域 |
「脳神経回路の形成・動作原理の解明と制御技術の創出」
(研究総括:小澤 瀞司 高崎健康福祉大学 健康福祉学部 教授) |
| 研究課題名 |
「可塑的神経回路を支えるシグナル伝達の分子基盤解明と制御」 |
| 研究代表者 |
尾藤 晴彦(東京大学 大学院医学系研究科 准教授) |
| 研究期間 |
平成22年10月~平成28年3月 |
JSTはこの領域で、脳神経回路の発生・発達・再生の分子・細胞メカニズムを解明し、さらに個々の脳領域で多様な構成要素により組み立てられた神経回路がどのように動作してそれぞれに特有な機能を発現するのか、それらの局所神経回路の活動の統合により、脳が極めて全体性の高いシステムをどのようにして実現するのかを追求します。またこれらの研究を基盤として、脳神経回路の形成過程と動作を制御する技術の創出を目指します。
上記研究課題では、可塑的神経回路を支えるシグナル伝達の分子基盤をシナプスレベル並びにシステムレベルで明らかにし、可塑的神経回路の脱構築・再構築を制御するための技術を開発します。
<研究の背景と経緯>
私たちは、日常で起こった出来事や学習によって得られた知識を、長い間覚えておくことができます。このような長期的な記憶は、脳の神経細胞同士の情報伝達の場であるシナプスの性質が長期的に変化することにより形成・保持されていると考えられています。例えば、あるシナプスに高頻度で入力が起こった後に、そのシナプスにおいて長期的に伝達効率が上昇する現象(長期増強)が知られています。このような長期的なシナプスの性質変化は、長期シナプス可塑性注2)と呼ばれます。神経細胞が状態を変化させる性質を持っていることが、それまでは記憶していなかった事柄を新しく記憶することを可能にするのです。これまでの研究により、シナプスの長期変化には、神経細胞の細胞体で新規遺伝子の発現が必要なことが明らかになっています。しかし、このような遺伝子の産物であるたんぱく質がどこのシナプスでどのように働くことにより、長期シナプス可塑性に関与しているのかについては不明でした(図1)。
<研究の内容>
本研究グループは、ラット脳から取り出して培養した細胞(初代神経培養細胞)や遺伝子改変マウスを材料にして、生化学的手法や蛍光ライブイメージング、大脳組織染色法などの多様な手法を組み合わせて解析を行いました。シナプス活動によって引き起こされた新規遺伝子の発現により、Arcと呼ばれるたんぱく質が合成されることはすでに知られていましたが、そのArcたんぱく質が細胞体からシナプスへ至る動態を世界で初めて解析しました。その結果、Arcたんぱく質は従来考えられていたように活動性の高いシナプスへ選択的に集積されるのではなく、逆に活動性の低いシナプスへ運ばれ集積されていることが明らかになりました。さらに、このArcのシナプス集積は別のシナプスたんぱく質であるカルシウムカルモジュリンキナーゼII注3)のβ型(CaMKIIβ)によって担われていることを明らかにしました。このCaMKIIβを介したArcの不活性シナプスへの集積度は、大脳の主要な興奮性の伝達に関わるAMPA型グルタミン酸受容体注4)のシナプス発現量と逆相関することも明らかになりました(図2)。これは、Arcの集積したシナプスは神経伝達物質に対する感度が低下する、つまり情報に対して“鈍く”なっていることを示唆します。
さらに、一部のシナプスのみに長期的な変化を引き起こした際のArcの働きを調べました。その結果、Arcは長期増強が起こっているシナプスを避けて、むしろ周りの活動性の低いと考えられるシナプスに集積するという現象が明らかになりました(図3)。つまり、増強されたシナプスはArcたんぱく質の影響を受けずに強化されたままで残り、一方、活動性の低いシナプスでは意図しないシナプス増強などを防ぐことができます。このようにシナプス間でメリハリをつけて、特定の記憶だけが長期記憶となることが分かりました。
これまで、脳の長期記憶に関するメカニズムとして、シナプスタグ方式注5)が知られていました。これは、長期的に情報を蓄えるべきシナプスを情報とは関係のないシナプスから見分けるための標識(タグ)が存在し、このタグがあるシナプスにだけ情報保持に必要なたんぱく質が集められていくという説です。今回は、シナプスタグ方式とは逆に情報を伝える必要のないシナプスにArcたんぱく質が集まることから、シナプスタグとは別の性質を持つタグ、すなわち「逆シナプスタグ」が存在すると考えられます。脳ではこのような巧妙な分子メカニズムによって選択的に長期記憶の形成が調節されていると考えられます。
<今後の展開>
Arcたんぱく質の不活性シナプスへの選択的集積は、これまで全く知られていなかった新しい動態制御の仕組み(図4)であり、シナプスタグ方式と合わせて、今後、記憶・学習の分子メカニズムを解明する上で基本的な概念となるものと思われます。
グルタミン酸受容体は大脳の主要な興奮性の伝達に関わるイオンチャネルであり、この機能や制御の破綻によりさまざまな神経・精神疾患が引き起こされることが最近の研究により明らかになってきています。今回発見したArcたんぱく質とCaMKIIβの相互作用はグルタミン酸受容体をシナプス部位において直接調節する分子機構であることから、この相互作用を標的とする新しいタイプの精神・神経治療薬の開発などにつながる可能性があります。
<参考図>
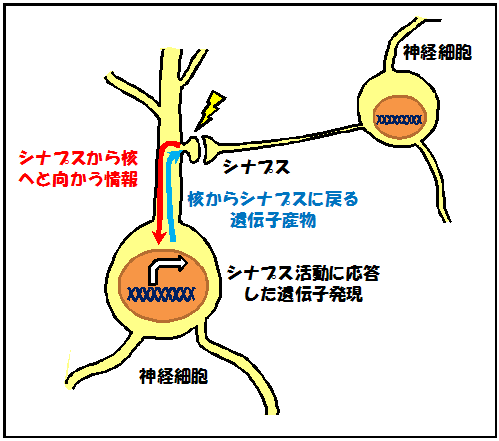
図1 シナプス活動によって引き起こされる遺伝子発現
長期記憶の形成には、神経細胞同士のつなぎ目であるシナプスの長期的な変化が必要と考えられています。また、このシナプスの長期変化には神経細胞の細胞体での新規遺伝子発現が必要なことが明らかになっています。しかしながら、遺伝子発現によってどのような変化が神経細胞やシナプスに起こるのかについては明らかになっていませんでした。
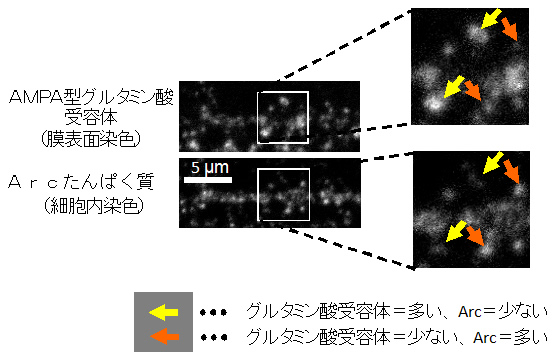
図2 シナプスに集積したArcはグルタミン酸受容体の量を減少させる
神経細胞の樹状突起でのArcたんぱく質の発現量とAMPA型グルタミン酸受容体の膜表面での発現量を、多重抗体蛍光染色法を使って比較したところ、シナプスでのArcの量が多い場合には膜表面にあるグルタミン酸受容体の量が少なく、逆に、Arcの量が少ない場合にはグルタミン酸受容体の膜発現量が多いことが明らかになりました。
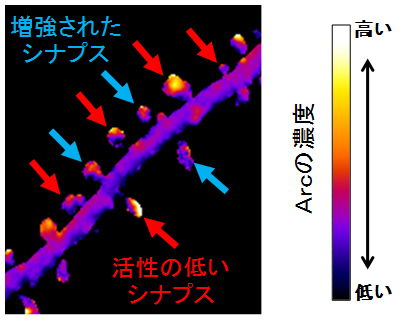
図3 長期増強刺激によって合成されたArcは増強されたシナプス以外のシナプスにより多く集まる
緑色蛍光たんぱく質GFPと融合したArcの動態を神経細胞の樹状突起にて観察したところ、長期増強刺激で発現誘導されたArcは強化されたシナプスにはあまり集積せず、むしろ、活動性の低いシナプスに集積することが明らかになりました。
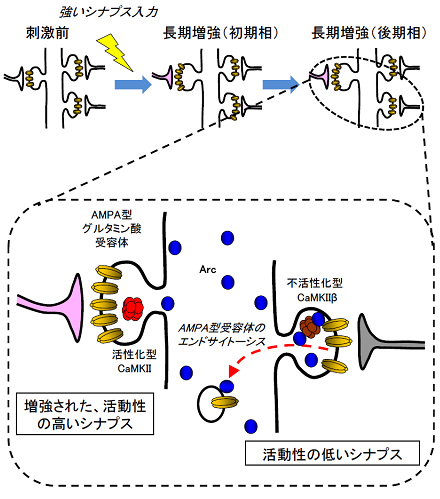
図4 Arcのシナプス集積とグルタミン酸受容体除去
長期増強を引き起こすような強いシナプス刺激を受けると、細胞体ではArcの発現誘導が起こります。Arcは不活性化型のCaMKIIβサブユニットに結合することにより、活動性の低いシナプスに選択的に集積します。この集積したArcはシナプス膜表面にあるAMPA型グルタミン酸受容体をエンドサイトーシスにより細胞内に取り込みます。このようなメカニズムがあれば、増強されたシナプスはArcの影響を受けずに強化されたままで残り、一方、活動性の低いシナプスは意図しないシナプス増強などを防ぐことができます。脳ではこのような巧妙な分子メカニズムによって長期記憶の形成が調節されていると考えられます。
<用語解説>
- 注1) シナプス
- 神経細胞と神経細胞との接合部および情報の伝達部位。送り手側の神経細胞はシナプス前部においてグルタミン酸などの神経伝達物質の放出し、受け手側の神経細胞はシナプス後部にある受容体などで神経伝達物質を受け取ることにより細胞間の情報伝達を行う。
- 注2) シナプス可塑性
- シナプス入力のパターンに応じて、神経伝達物質の放出および受け取りなどの確立や様式が変化することにより、細胞間の神経伝達効率が変化すること。その持続時間に応じて短期相・長期相などに細分される。
- 注3) カルシウムカルモジュリンキナーゼII
- カルシウムとカルモジュリンが結合することによりキナーゼ活性が上昇するキナーゼファミリーの1つ。いくつかのサブタイプが存在するが、神経細胞ではα型とβ型が主要な役割を担っていると考えられている。
- 注4) AMPA型グルタミン酸受容体
- シナプス後部の膜表面においてグルタミン酸を受け取るイオンチャネル型の受容体の1つ。神経細胞における細胞興奮性活動を引き起こす主要な構成因子である。
- 注5) シナプスタグ方式
- 強い刺激を受けたシナプスの伝達効率を選択的に増強する方式の1つ。刺激を受けたシナプスはシナプスタグによって標識される。シナプス増強に必須なたんぱく質は標識されたシナプスに補足され機能することにより、シナプス選択的な変化が引き起こされる。
<論文名>
“Inverse synaptic tagging of inactive synapses via dynamic interaction of Arc/Arg3.1 with CaMKIIβ”
(Arc/Arg3.1とCaMKIIβの動的相互作用による不活性シナプスへの逆シナプスタギング)
doi: 10.1016/j.cell.2012.02.062
<お問い合わせ先>
<研究に関すること>
尾藤 晴彦(ビトウ ハルヒコ)
東京大学 大学院医学系研究科 准教授
〒113-0033 東京都文京区本郷7-3-1
Tel:03-5841-3560 Fax:03-3814-8154
E-mail:
<JSTの事業に関すること>
石井 哲也(イシイ テツヤ)
科学技術振興機構 戦略研究推進部
〒102-0076 東京都千代田区五番町7 K’s五番町
Tel:03-3512-3531 Fax:03-3222-2066
E-mail: