■ ポイント ■
- X線結晶構造解析手法と機能解析手法を用い、ウイルスRNA合成酵素と宿主翻訳因子が形成
する複合体によるRNA合成過程を詳細に解析
- 宿主翻訳因子がRNAウイルスのRNA合成の伸長反応を促進する別機能を有することを発見
- 太古では翻訳因子が元来RNA合成の因子としての機能を有していた可能性を示唆
■ 概 要 ■
独立行政法人 産業技術総合研究所【理事長 野間口 有】(以下「産総研」という)バイオメディカル研究部門【研究部門長 織田 雅直】RNAプロセシング研究グループ 富田 耕造 研究グループ長、竹下 大二郎 研究員らは、Qβウイルス注1)由来のRNA注2)合成酵素(β-サブユニット)と宿主注3)(大腸菌)由来の2つの翻訳因子注4)が複合体を形成し、複合体中の翻訳因子がRNAの合成伸長過程を促進する新たな機能を有することを発見した。この発見は、RNAをゲノム注5)として有していたと考えられる太古生命体では、翻訳因子がRNA合成酵素の補因子としての役割を担っていた可能性を示唆する。
本研究は独立行政法人 科学技術振興機構(JST) 戦略的創造研究推進事業 個人型研究(さきがけ)「RNAと生体機能」研究領域【研究総括:微生物化学研究所長 野本 明男】における研究課題「RNA末端合成プロセス装置の分子基盤」、独立行政法人 日本学術振興会(JSPS) 最先端次世代開発支援プログラム(NEXT Program)「RNA合成酵素の反応制御機構の解明(代表者:富田 耕造)」の一環として行われた。また、X線回折データは大学共同利用機関法人 高エネルギー加速器研究機構(KEK)のフォトンファクトリーを利用して取得した。本成果は、2012年1月16日(日本時間)に、「 Nature Structural & Molecular Biology 」のオンライン版に掲載される。
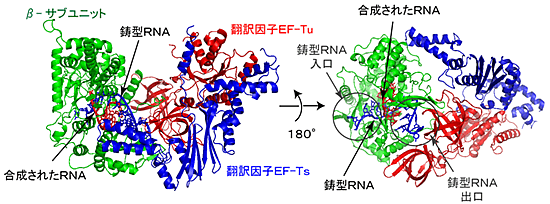
RNA合成伸長過程のQβウイルスRNA合成酵素-翻訳因子複合体のRNA合成伸長過程の構造
- RNA合成酵素:β-サブユニット(緑)、翻訳因子EF-Tu(赤)、EF-Ts(青)、鋳型RNA(スティック表示:青)、合成されたRNA(スティック表示:赤)
<研究の社会的背景>
RNAウイルスのゲノムの宿主内での複製注6)や転写注6)のプロセスの大部分は宿主由来のタンパク質を必要とするが、複製、転写の際のRNA合成はウイルス自身が有するRNA合成酵素がその触媒活性を担っている。一部のRNAウイルスでは、RNAゲノムの複製、転写を担うウイルス由来のRNA合成酵素が、宿主の翻訳因子と複合体を形成し、その複合体形成がRNAウイルスのゲノムの転写や複製に必須であることが知られている。ウイルス由来のRNA合成酵素と宿主の翻訳因子との複合体形成やRNAウイルスのゲノムの宿主内での複製、転写における宿主翻訳因子の役割を分子レベルで解明することは、新たな抗ウイルス剤の開発の基盤につながると期待されている。また、現存するほとんどの生物では、RNAを合成するシステムとタンパク質を合成するシステムは互いに独立したプロセスであるが、タンパク質合成に必要な因子がRNA合成にも必要な因子として組み込まれているウイルスのRNA合成酵素複合体の研究は、RNA合成システムがタンパク質合成システムへと進化した分子レベルでの仕組みや、生命進化における翻訳因子の起源について、学術的に重要な知見が得られる可能性を秘めている。
<研究の経緯>
産総研では、RNA合成酵素群の研究を行っており、これまで、鋳型を用いないでRNAを合成する酵素群やウイルス由来の鋳型を用いてRNAを合成する酵素の機能、構造、進化、制御機構の研究を推進してきた。さらに、核酸性の鋳型注7)を用いてRNAを合成するウイルス由来のRNA合成酵素のうち、大腸菌に感染するQβウイルス由来のRNA合成酵素とQβウイルスに感染した細胞内の翻訳因子との複合体形成機構を解明した(2010年8月24日 産総研・JST共同プレス発表)。今回、このRNA合成酵素と翻訳因子の複合体が、RNAの合成を開始しRNA鎖を伸長、合成していく過程についてX線構造解析、また構造をもとにした機能解析を行い、翻訳因子のRNA合成過程における役割の解明を目指した。なお、X線回折データはKEKのフォトンファクトリー構造生物学ビームラインBL-17Aを利用して取得した。
<研究の内容>
翻訳因子であるEF-Tu、EF-Ts注8)は全生物において普遍的に存在し、タンパク質合成における伸長サイクルに必要不可欠なタンパク質である。1970年代の初頭に、翻訳因子EF-Tu、EF-Tsが大腸菌に感染するQβウイルスのQβ複製酵素複合体注9)を形成する不可欠なサブユニットであることが報告された。この報告は翻訳因子の別の役割、機能を示す最初のものであった。
その後、いくつかの植物、動物、他のバクテリアに感染するRNAウイルスのゲノムにコードされているRNA合成酵素が宿主由来の翻訳因子と相互作用しうることが報告され、その相互作用がRNAウイルスのゲノムの複製、転写に必要であることが報告されてきた。しかしながら、長年、ウイルスにコードされているRNA合成酵素が宿主由来の翻訳因子と相互作用し、複合体を形成する分子機構や、複合体中の翻訳因子のタンパク質合成での機能とは異なったRNA複製、転写における役割については謎であり、その分子機構は明らかにされていなかった。
Qβウイルスは、1本鎖RNAをゲノムとしてもつ、大腸菌に感染するウイルスであり、Qβ複製酵素複合体によってそのRNAゲノムの複製、転写を行う。Qβ複製酵素複合体はウイルス由来のRNA合成酵素であるβサブユニット、宿主由来の翻訳因子EF-Tu、EF-Tsとリボソームタンパク質S1注10)から構成される。とくにRNAの複製、転写にはβサブユニットとEF-Tu、EF-Tsとが3者複合体を形成することが必要である。本研究では、この3者複合体がRNA合成を開始し、RNA鎖が伸長していく過程の複数の構造について、X線結晶構造解析と構造に基づいた生化学機能解析を行った。
その結果、以下の1)~5)のことが判明した。
- 1)RNA合成開始時には、鋳型となるRNAの3´末端のアデニン(A)注11)はRNA合成の鋳型となるのではなく、RNA合成の最初のヌクレオチド(GTP)注2)のグアニン(G)注11)と鋳型RNAの3´末端から2番目のシトシン(C)注11)との塩基対と相互作用して、開始時の複合体を安定化させ、RNA合成開始を効率よく行わせる役割を担っている(図1)。
- 2)RNA合成伸長過程では、8ヌクレオチドの長さのRNAが合成されるまで、合成されたRNAは鋳型RNAと2重鎖を形成し、その2重鎖は複合体中のEF-Tu、EF-Tsの方向へ進行する。この方向は、鋳型RNAのリン酸骨格がEF-Tuと水素結合を形成することによって決定される(図2)。
- 3)9ヌクレオチドの長さのRNAが合成されると、鋳型RNAの3´末端の突出したアデニンはβサブユニットのC末端領域(クサビ領域)と相互作用する。その結果、鋳型RNAと合成されたRNAの間の水素結合は不安定になる(図3左)。
- 4)10ヌクレオチドの長さのRNAが合成されると、鋳型RNAと合成されたRNAからなる2重鎖はさらに複合体中のEF-Tu、EF-Tsの方向へ進行し、鋳型RNAの3´末端の突出したアデニンはクサビ領域やEF-Tuと相互作用することによって反転し、βサブユニットとEF-Tuの間に形成されるトンネル(鋳型RNA出口)の方向へ導かれる(図3中央)。また、鋳型RNAと合成されたRNAからなる2重鎖RNA間の水素結合はさらに不安定になる。
- 5)さらに、RNAが伸長すると(14ヌクレオチド)、鋳型RNAと合成されたRNAの間の水素結合はクサビ領域によってほとんど解かれ、鋳型RNAは鋳型RNA出口トンネルへ入り込み、また合成されたRNAの5´側は複合体から解離する(図3右)。
以上の結果から、ウイルスのRNA合成酵素と複合体を形成する宿主由来の翻訳因子は、RNA合成伸長過程において、鋳型RNAと合成されたRNAの2重鎖をほどき、効率よくRNA伸長合成が行われるのを補助するとともに、鋳型RNAの出口トンネルをRNA合成酵素と共同で形成することによって、RNAウイルスのゲノムの転写、複製が完了するまで、鋳型RNAが複合体から解離してしまうのを防ぐ役割もあることがわかった。
RNA分子が化学反応を触媒する機能を有していることが発見され、太古生命体では、生命体を構成する分子はRNAであり、RNA分子が遺伝情報の保存分子であり、かつ化学反応を触媒する酵素であったとする「RNAワールド仮説」が提唱されている。長い年月を経て、RNAの遺伝情報の保存分子としての役割が化学的に安定なDNAへ、そして、RNAの酵素活性の役割がタンパク質へと置き換わり、現在の生命になったと考えられている。また、生命の進化において、RNA合成-複製システムはタンパク質合成システムよりも先に出現したと考えられている。これらの仮説や考えは多くの研究により支持されている。今回の研究から明らかになった翻訳因子にRNA合成、伸長を促進する役割があるという事実は、RNAゲノムからなる太古生命体では、翻訳因子は元来、RNAゲノムの複製や転写を促進する補因子としての役割を担っており、その後出現した現在のタンパク質合成システムが、このRNA合成補因子を翻訳因子として取り込んだ可能性を示していると考えられる。
<今後の予定>
ウイルスのRNA合成酵素の中には、宿主由来の翻訳因子以外のタンパク質合成に関わる因子と複合体を形成することも知られている。これらの因子のRNA合成における機能の解明を行う。また、これらの解析から、生命が進化する過程でRNAの機能がタンパク質へ置き替わる遷移の分子機構、分子進化、RNA合成システム、タンパク質合成システムの進化、起源を明らかにしていく。
<参考図>
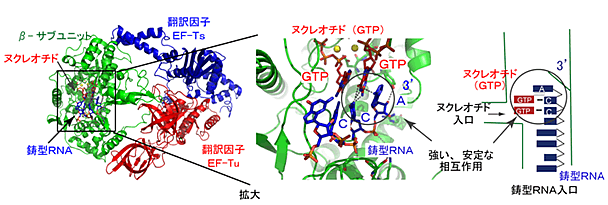
図1 RNA合成開始時のQβウイルスRNA合成酵素(βサブユニット:緑)と翻訳因子EF-Tu(赤)、EF-Ts(青)との複合体
鋳型RNA(スティック表示:青)、ヌクレオチド(GTP:スティック表示:赤)。鋳型RNAの末端のAはGTPと鋳型のCとの塩基対と相互作用し、RNA合成複合体を安定化する。
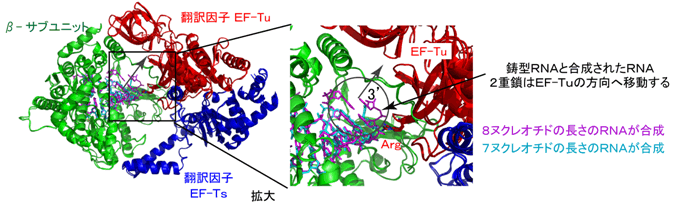
図2 QβウイルスRNA合成酵素複合体によるRNA伸長過程
合成された7ヌクレオチドRNAと鋳型RNAの2本鎖(シアン)、合成された8ヌクレオチドRNAと鋳型RNAの2本鎖(マゼンタ)。
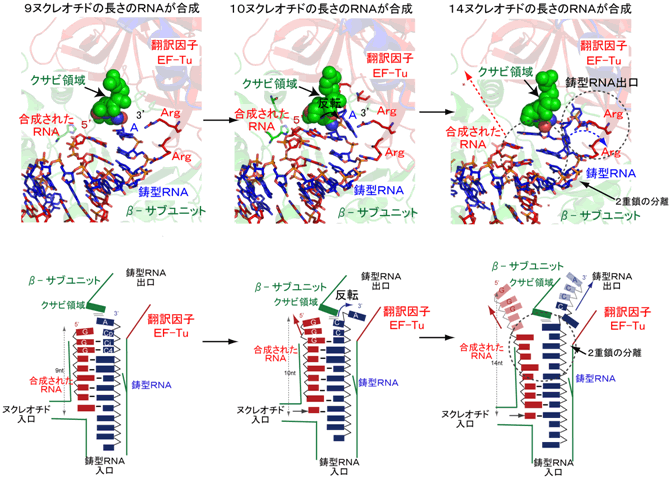
図3 RNA伸長過程のQβウイルスRNA合成酵素複合体の構造
9ヌクレオチド(左)、10ヌクレオチド(中央)、14ヌクレオチド(右)の長さのRNAが合成された状態。
<用語解説>
- 注1) Qβウイルス
- 1本鎖RNAをゲノムとして有し、大腸菌に感染して増殖するウイルスの一種。このウイルスが感染すると、大腸菌は溶菌し、死滅する。
- 注2) RNA、ヌクレオチド(GTP)
- 核酸にはDNAとRNAが存在する。DNAはA、G、C、Tの4種類の構成要素(ヌクレオチド)が連結したものであり、一方RNAはA、G、C、Uの4種類のヌクレオチドが連結したものである。
- 注3) 宿主
- 一般に、ウイルスは自分自身だけでは増殖できず、他の生物に感染して、その生物のもつタンパク質などの助けを借りて増殖する。この感染先の生物、細胞のことを宿主(しゅくしゅ)と呼ぶ。
- 注4) 翻訳因子
- 生体内でのタンパク質の合成を翻訳という。タンパク質はDNA(あるいはRNA)の配列(遺伝暗号)をもとにして、アミノ酸が連結されたタンパク質へと変換される。翻訳因子は、タンパク質の翻訳過程に関与するタンパク質のことをいう。
- 注5) ゲノム
- 生物のもつ遺伝情報が保存されている分子。通常の生物のゲノムはDNAであるが、ウイルスには、RNAをゲノムとして有するものがある。
- 注6) 複製、転写
- ある核酸(DNA、RNA)の配列があるとき、その配列と全く同じ配列の核酸を合成することを複製という。また、ある核酸(DNA、RNA)の配列の相補的な配列を合成することを転写という。
- 注7) 核酸性の鋳型
- 生体内での核酸(DNAやRNA)の合成は通常、その設計図となる核酸配列が存在し、その配列と塩基が対になる配列(相補性:A⇔U(T)、C⇔G)の塩基が並ぶようにDNAやRNAの合成が行われる。その完成予定の配列の対となる配列を鋳型と呼び、合成される配列は鋳型と相補的である。
- 注8) EF-Tu、EF-Ts
- 全ての生物に存在する翻訳伸長因子である。EFはElongation Factorの略。EF-Tuはアミノ酸の結合したtRNAをリボソームと呼ばれるタンパク質合成装置へ運搬し、EF-TsはEF-Tuをリサイクルする役割を果たしている。いずれもタンパク質の合成システムに必須のタンパク質である(下図参照)。
- 注9) Qβ複製酵素複合体
- QβウイルスゲノムRNAの複製や転写を行う酵素複合体。
- 注10) リボソームタンパク質S1
- 生体内でのタンパク質合成はリボソームと呼ばれる複合体で行われる。リボソームは数十種類のタンパク質と3つのRNAからなるが、リボソームタンパク質S1はリボソームを構成するタンパク質の一種。
- 注11) アデニン(A)、グアニン(G)、シトシン(C)
- ヌクレオチドに含まれる塩基。塩基部分がヌクレオチドの特異性を決定する。RNAは4種類のヌクレオチドからなるが、その塩基部分はグアニン(G)、アデニン(A)、シトシン(C)、ウラシル(U)である。DNAの場合、グアニン(G)、アデニン(A)、シトシン(C)、チミン(T) の4種類の塩基がある。
<お問い合わせ先>
<研究内容に関すること>
富田 耕造(トミタ コウゾウ)
産業技術総合研究所 バイオメディカル研究部門 RNAプロセシング研究グループ 研究グループ長
〒305-8566 茨城県つくば市東1-1-1 中央第6-13
Tel:029-861-6085
E-mail:
原口 亮治(ハラグチ リョウジ)、木村 文治(キムラ フミハル)
科学技術振興機構 イノベーション推進本部 研究推進部(研究促進担当)
〒102-0076 東京都千代田区五番町7 K’S五番町
Tel:03-3512-3525 Fax:03-3222-2067
E-mail:
<プレス発表/取材に関する窓口>
産業技術総合研究所 広報部 報道室
〒305-8568 茨城県つくば市梅園1-1-1 中央第2 つくば本部・情報技術共同研究棟8F
Tel:029-862-6216 Fax:029-862-6212
E-mail:
科学技術振興機構 広報ポータル部
〒102-8666 東京都千代田区四番町5-3
Tel:03-5214-8404 Fax:03-5214-8432
E-mail: