JST 課題達成型基礎研究の一環として、東京大学 大学院薬学系研究科の田口 友彦 特任准教授と新井 洋由 教授らは、コレラ菌が放出し、コレラを発症する原因となる「コレラ毒素注1)」が細胞内に運ばれる経路を発見し、その運搬経路が毒素感染に必須であることを明らかにしました。
これまで、コレラ毒素は細胞外から細胞内に取り込まれてその毒性を発揮することが分かっていましたが、どのようにして毒性を発揮する場所まで運ばれるのか、詳細な細胞内の運搬経路は明らかになっていませんでした。運搬経路を知ることは、コレラの細胞毒性発現を抑制するための1つの有力なアプローチとして考えられ、興味が持たれていました。
今回研究グループは、生体内のほとんどの臓器、組織に存在している「エベクチン2たんぱく質注2)」の機能を研究する中で、エベクチン2たんぱく質がコレラ毒素の細胞内での運搬経路のうち、これまで解明されていなかった部分(細胞膜からゴルジ体にいたる過程)の物質輸送を制御するたんぱく質であることを発見しました。エベクチン2たんぱく質のない細胞では、コレラ毒素が細胞内の運搬経路の途中で蓄積してしまい、毒素感染の指標の上昇も抑制されました。
また、エベクチン2たんぱく質は、ホスファチジルセリンと呼ばれる「リン脂質注3)」の一種と結合し運搬経路を制御することから、細胞内物質輸送におけるリン脂質の重要性も明らかとなりました。
今回の発見により、コレラ毒素の細胞への侵入経路が明らかとなり、この運搬経路を標的とした新たな抗毒素薬の開発などにつながるものと期待されます。
本研究成果は、米国科学アカデミー紀要「Proceedings of the National Academy of Sciences of the United States of America」のオンライン速報版で2011年9月12日の週(米国東部時間)に公開されます。
本成果は、以下の事業・研究領域・研究課題によって得られました。
戦略的創造研究推進事業 チーム型研究(CREST)
| 研究領域 |
「代謝調節機構解析に基づく細胞機能制御基盤技術」
(研究総括:西島 正弘 昭和薬科大学 特任教授) |
| 研究課題名 |
「生体膜リン脂質多様性の構築機構の解明と高度不飽和脂肪酸要求性蛋白質の同定」 |
| 研究代表者 |
新井 洋由(東京大学 大学院薬学系研究科 教授) |
| 研究期間 |
平成18年10月~平成24年3月 |
JSTはこの領域で、細胞内の代謝変化を統合的あるいは網羅的に解析し、細胞機能の制御メカニズムや恒常性維持のメカニズムを明らかにし、細胞機能を効率的に制御・変換したり、恒常性の乱れを改善・回復させる細胞制御基盤技術の創出を目指しています。上記研究課題では、リン脂質分子種多様性形成に関わる分子群の同定、および高度不飽和脂肪酸鎖を持つリン脂質の生物的意義の解明に焦点をしぼり、生体膜機能の本質的問題の解決を目指します。
<研究の背景と経緯>
コレラは、コレラ菌(Vibrio cholerae)を病原体とする経口感染症であり、小腸に到達したコレラ菌が放出するコレラ毒素によって、腸の表面にある細胞の機能が損なわれることによって発症します。コレラの感染力は大変強く、過去に幾度も世界的大流行を引き起こし、現在もコンゴ、ハイチなどではコレラ感染によって多くの人が死亡しています。
これまでに、コレラ毒素が腸の表面にある細胞に取り込まれ、の細胞小器官注4)であるゴルジ体や小胞体を通過しながら最終的にサイトゾル(細胞内の部分の呼称で、細胞質から細胞内小器官を除いた部分)という空間に侵入することによって毒性を発揮することは明らかになっていました。しかし、どのように細胞膜からゴルジ体へ運搬されるのか分かっていませんでした(図1)。
<研究の内容>
研究グループは今回、アフリカミドリザルの細胞(COS-1細胞)が秩序だった細胞小器官の分布を持つという田口特任准教授らの発見(参考論文参照:2007年のCRESTによる成果、図2)を利用することで、コレラ毒素の細胞内運搬経路を明らかにし、その運搬経路を制御する分子の同定を行う研究を開始しました。
その結果、以下の4点が明らかになりました。
- 1)コレラ毒素は、細胞膜から取り込まれた後、細胞膜へ再回収(リサイクル)する経路に関わる細胞小器官のリサイクリングエンドソーム注5)を経由してゴルジ体に運搬されること(図3)
- 2)リサイクリングエンドソームに局在するエベクチン2たんぱく質が、コレラ毒素のリサイクリングエンドソームからゴルジ体への運搬を制御すること(図4)
- 3)リサイクリングエンドソームには、ホスファチジルセリンというリン脂質が豊富に存在すること(図5)
- 4)ホスファチジルセリンと結合するたんぱく質(LactC2)を細胞内で過剰発現することによって、コレラ毒素のゴルジ体への輸送が抑制されること(図6)
本研究によって、リサイクリングエンドソームの細胞内物質輸送における新しい機能(リサイクリングエンドソームからゴルジ体への輸送)と、リサイクリングエンドソームの重要な構成因子(エベクチン2たんぱく質、ホスファチジルセリン)が明らかになりました。リサイクリングエンドソームから細胞膜への物質輸送はすでに知られていましたが、リサイクリングエンドソームからゴルジ体への物質輸送は、今回初めて発見されたものです。サイトゾルへ輸送されることで毒性を発揮するほかの毒性たんぱく質(赤痢菌が放出するシガ毒素、毒素原性大腸菌が放出する易熱性エンテロトキシン、ヒマの種子から単離されるリシンなど)もリサイクリングエンドソームを介する輸送経路によってサイトゾルへ運ばれ、細胞毒性を発揮している可能性が出てきました。
<今後の展開>
今回の発見によって、コレラ毒素のサイトゾルへの侵入をリサイクリングエンドソームの段階で阻害する薬剤開発を行うことが可能になり、毒素の持つ酵素活性を阻害する薬剤の開発だけではなく、物質輸送の視点に立脚した抗毒素薬の開発という新しい治療戦略を提唱できました。
また、リサイクリングエンドソームからゴルジ体への物質輸送経路は、毒素のためだけに細胞に用意されているとは考えにくく、細胞が通常この運搬経路によって輸送しているたんぱく質を同定することで、リサイクリングエンドソームの新たな細胞生物学的機能が明らかになることが期待されます。
<参考図>
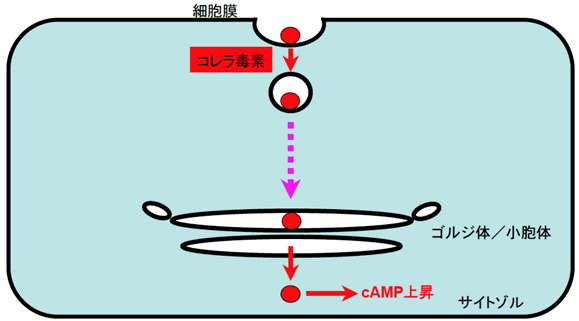
図1 コレラ毒素の細胞内侵入経路
コレラ菌によって放出されたコレラ毒素(●)は、小腸上皮細胞膜に発現しているGM1糖脂質に結合し、細胞内へ取り込まれます。取り込まれたコレラ毒素は、ゴルジ体や小胞体を経由して最終的にサイトゾル(青色の部分)へ脱出し、環状アデノシン一リン酸(cAMP)濃度の上昇を引き起こします。細胞膜からゴルジ体へ至る輸送経路については不明でした。
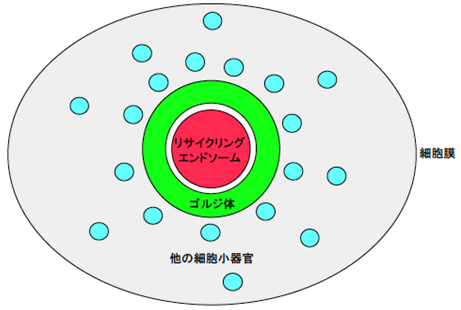
図2 COS-1細胞における細胞小器官の空間分布
COS-1細胞では、ゴルジ体はリング状の構造をとり、その内側にリサイクリングエンドソームが存在します。この特徴がリサイクリングエンドソームの顕微鏡による観察を容易にしました。ほかの細胞小器官はゴルジ体のリング構造の外側に分布します。
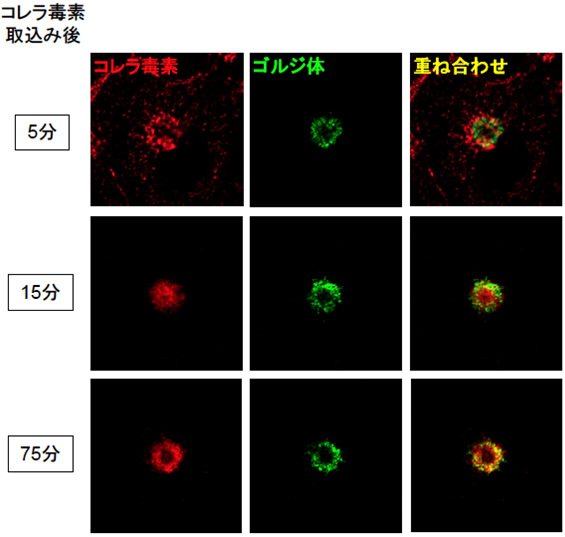
図3 細胞に侵入した後のコレラ毒素の移動経路の可視化
蛍光標識したコレラ毒素をCOS-1細胞に取り込ませ、取り込ませた時間を0分として一定時間経過後、蛍光顕微鏡で観察しました。ゴルジ体は、緑色で染色しています。コレラ毒素が、15分後にリサイクリングエンドソームへ(ゴルジ体の形作るリング構造の内側)、75分後にゴルジ体へ運搬されるのが容易に見て取れます。
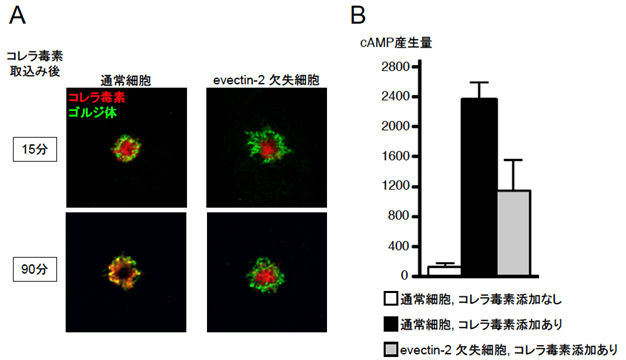
図4 エベクチン2たんぱく質欠失によるコレラ毒素の輸送遅延
- (A)エベクチン2たんぱく質を欠失させた細胞では、毒素はリサイクリングエンドソームまでは到達できますが(15分後)、90分後もリサイクリングエンドソームに蓄積したままでゴルジ体に到達できていないことが観察されます。一方、通常細胞では90分後には、毒素のほとんどがゴルジ体に到達しているのが観察されます。
- (B)エベクチン2たんぱく質を欠失させた細胞では、cAMP濃度の上昇が、通常細胞(コレラ毒素添加有り)と比較して抑制されているのが分かります。
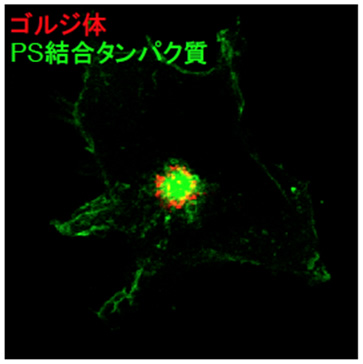
図5 リサイクリングエンドソーム中のホスファチジルセリンの可視化
ホスファチジルセリン(PS)と特異的に結合するたんぱく質(LactC2)に蛍光たんぱく質(GFP)をつなげたもの(図中:PS結合たんぱく質)をCOS-1細胞で発現させました。ゴルジ体は赤色で染色しています。LactC2たんぱく質がゴルジ体の内側(リサイクリングエンドソーム)に集積していることが観察されることから、ホスファチジルセリンがリサイクリングエンドソームに豊富にあることが分かりました。
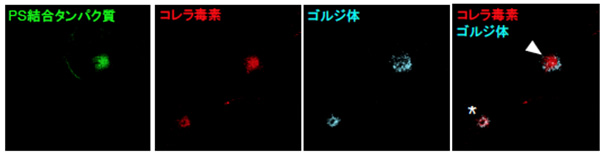
図6 ホスファチジルセリンに結合するたんぱく質の過剰発現によるコレラ毒素の輸送遅延
ホスファチジルセリン(PS)に特異的に結合するたんぱく質(LactC2たんぱく質)をCOS-1細胞で過剰に発現させ、その後、コレラ毒素の取り込みを行いました。図には2つの細胞があり、矢頭で示しているのがLactC2たんぱく質を発現している細胞、*で示しているのが通常細胞です。通常細胞では、コレラ毒素がゴルジ体に到達していますが、LactC2たんぱく質を過剰に発現している細胞では、ゴルジ体に到達できずリサイクリングエンドソームで蓄積してしまっていることが分かります。つまり、LactC2たんぱく質が過剰に存在するためにエベクチン2たんぱく質がホスファチジルセリンと結合できなくなり、コレラ毒素が運ばれにくくなったと考えられます。
<用語解説>
- 注1) コレラ毒素
- コレラは激しい水様性の下痢を伴う致死的な細菌性感染症。コレラは、飲食物などに含まれるコレラ菌が経口感染し、腸管においてコレラ菌がコレラ毒素を放出することにより引き起こされる。コレラ毒素は、1つのAサブユニットと5つのBサブユニットから構成されるたんぱく質性の毒素であり、AサブユニットがADP-リボシル化能を持つ酵素である。コレラ毒素は腸管の上皮細胞に取り込まれると、最終的に上皮細胞のサイトゾルへ運搬され、次いで3量体Gたんぱく質のADP-リボシル化を行うことで、3量体Gタンパク質の恒常的な活性化を引き起こす。その結果として環状アデノシン一リン酸(cAMP:cyclic adenosine monophosphate)の濃度が異常に上昇し、細胞、組織から電解質と水が大量に漏出することで、コレラ特有の激しい下痢が起きると考えられている。
- 注2) エベクチン2(evectin-2)たんぱく質
- 1999年に発見されたたんぱく質。生体内のほとんどの臓器、組織に存在しており、細胞内ではゴルジ体の近縁部に存在することが分かっていたが、その機能についてはこれまで解析されていなかった。
- 注3) リン脂質
- 構造中にリン酸エステル結合を持つ脂質の総称。両親媒性を持ち、脂質二重膜を形成して糖脂質やコレステロールとともに生体膜の主要な構成成分となるほか、生体内でのシグナル伝達にも関わる。リン脂質は、大きく分けてグリセロールを骨格とするグリセロリン脂質と、スフィンゴシンを骨格とするスフィンゴリン脂質の2つがあり、ホスファチジルセリンはグリセロリン脂質に属する。
- 注4) 細胞小器官
- 細胞の内部で特に分化した形態を持つ構造の総称。一般的に膜で囲まれた構造を指す。小胞体(たんぱく質の合成/糖の付加)、ゴルジ体(糖鎖修飾)、リソソーム(たんぱく質の分解)、エンドソーム(注5参照)、ミトコンドリア(エネルギー産生)など、それぞれ固有の機能を持っている。
- 注5) リサイクリングエンドソーム
- 細胞が細胞外の物質を取り込む営みをエンドサイトーシスと呼び、エンドサイトーシスによって取り込まれた物質が運ばれる細胞小器官を総称してエンドソームと呼ぶ。リサイクリングエンドソームは、エンドソームの一種であり、エンドサイトーシスで取り込まれた物質が細胞膜へ再回収する過程(リサイクル)で通過するエンドソームのことをいう。
<論文タイトルと著者>
“Intracellular phosphatidylserine is essential for retrograde membrane traffic through endosomes”
(細胞内のホスファチジルセリンはエンドソームを通過する逆行性膜輸送に必須である)
doi: 10.1073/pnas.1109101108
<参考論文名および著者名>
“Spatial segregation of degradation- and recycling-trafficking pathways in COS-1 cells”
(COS-1細胞における分解輸送経路と再回収輸送経路の空間的分離)
Misaki, R., Nakagawa, T., Fukuda, M., Taniguchi, N., and Taguchi, T., Biochem Biophys Res Commun 360, 580-585 (2007).
<お問い合わせ先>
<研究に関すること>
田口 友彦(タグチ トモヒコ)
東京大学 大学院薬学系研究科 特任准教授
〒113-0033 東京都文京区本郷7-3-1
Tel:03-5841-4721
E-mail:
新井 洋由(アライ ヒロユキ)
東京大学 大学院薬学系研究科 教授
〒113-0033 東京都文京区本郷7-3-1
Tel:03-5841-4720
E-mail:
<JSTの事業に関すること>
石井 哲也(イシイ テツヤ)
科学技術振興機構 イノベーション推進本部 研究領域総合運営部
〒102-0075 東京都千代田区三番町5 三番町ビル
Tel:03-3512-3524 Fax:03-3222-2064
E-mail: