JST 課題解決型基礎研究の一環として、大阪バイオサイエンス研究所の古川 貴久 研究部長らは、マウスを使った研究で、網膜色素変性症注1)を引き起こす新しい仕組みを発見しました。
網膜色素変性症は、眼の中で光を神経の信号に変える働きをする網膜に異常をきたす遺伝性、進行性の難病です。眼の網膜の最外層にあり、光センサーの役割をする視細胞注2)が細胞死することにより、失明や重篤な視力障害を引き起こしますが、その根本的な治療法はいまだに確立されていません。
本研究グループは、特定の遺伝子を働かないようにしたノックアウトマウスを用いた実験により、視細胞に発現するリン酸化酵素(セリンスレオニンキナーゼ)Makが、網膜色素変性症の原因遺伝子の1つとして知られているRP1たんぱく質の活性をコントロールすることで、光をキャッチするアンテナの役割を担う繊毛注3)の長さ調節を行っていることを発見しました。また、この仕組みが壊れると繊毛が伸びすぎて長くなり、これにより視物質注4)が細胞体に蓄積して視細胞の細胞死が引き起こされ視覚障害が生じ、網膜色素変性症につながることが分かりました。
これは網膜色素変性症を引き起こす新しいメカニズムの解明であり、網膜色素変性症の診断法や治療法の開発に向けて一歩前進したと言えます。
本研究成果は、米国科学雑誌「米国科学アカデミー紀要(PNAS)」のオンライン速報版で2010年12月6日の週(米国東部時間)に公開されます。
本成果は、以下の事業・研究領域・研究課題によって得られました。
戦略的創造研究推進事業 チーム型研究(CREST)
| 研究領域 |
: |
「脳神経回路の形成・動作原理の解明と制御技術の創出」
(研究総括:小澤 瀞司 高崎健康福祉大学 健康福祉学部 教授)
|
| 研究課題名 |
: |
「網膜神経回路のシナプス形成と生理機能発現の解析」 |
| 研究代表者 |
: |
古川 貴久(大阪バイオサイエンス研究所 発生生物学部門 研究部長) |
| 研究期間 |
: |
平成21年11月~平成27年3月 |
JSTはこの領域で、脳神経回路の発生・発達・再生の分子・細胞メカニズムを解明し、さらに個々の脳領域で多様な構成要素により組み立てられた神経回路がどのように動作してそれぞれに特有な機能を発現するのか、それらの局所神経回路の活動の統合により、脳が極めて全体性の高いシステムをどのようにして実現するのかを追求します。またこれらの研究を基盤として、脳神経回路の形成過程と動作を制御する技術の創出を目指します。
上記研究課題では、網膜に注目してシナプスが特異的に結合する分子メカニズムおよび網膜神経回路の生理機能と動作メカニズムの解明を目的とし、網膜神経回路がどのようなメカニズムと機能原理に基づき視覚情報処理を行っているかを明らかにします。
<研究の背景と経緯>
網膜色素変性症(retinitis pigmentosa)は、視細胞が変性し細胞死を引き起こすことにより、失明や重篤な視覚障害を引き起こす疾患です。緑内障、糖尿病網膜症とともに、中途失明の3大原因の1つです。その根本的な治療法は確立されておらず、厚生労働省から難病指定を受けています。網膜色素変性症の代表的な原因遺伝子の1つにRP1(retinitis pigmentosa 1)がありますが、RP1が網膜色素変性症を引き起こす仕組みは明らかになっていませんでした。
また眼の網膜には、視細胞と呼ばれる神経細胞がシート状に配列し、光センサーの役割を果たしています(図1)。視細胞には、光をキャッチするアンテナの役割を果たすために繊毛が形成されます。繊毛はさまざまな細胞に形成され、外部情報をキャッチするためのアンテナとして機能することが知られており、繊毛の長さは細胞ごとに決まっています(図2)。しかし、この繊毛が決まった長さに作られる仕組みはこれまでよく分かっていませんでした。
<研究の内容>
本研究グループは今回、網膜の視細胞に特異的に発現する遺伝子のスクリーニング注5)の中から視細胞に強く発現するリン酸化酵素(セリンスレオニンキナーゼ)Makに注目し、Mak遺伝子を欠損したマウスを観察したところ、人間の網膜色素変性症に似た症状が見られることを発見しました。このマウスでは、生後1ヵ月から進行性に視細胞が死んでいく変性が観察され、光を感知する能力が劣っていることが電気生理学的な測定から分かりました。そこで網膜の組織を詳細に観察したところ、このMak欠損マウスでは視細胞の繊毛と呼ばれる神経細胞のアンテナの部分が異常に長くなっており、視物質であるロドプシンが細胞体に蓄積していました(図3)。これらより、繊毛の適切な長さ調節が視細胞の生存に必須であることが分かりました(図4)。さらに、繊毛においてMakと共に局在が一致するRP1たんぱく質に注目し、培養細胞を用いた繊毛の形成実験により、このMakが網膜色素変性症の原因遺伝子であるRP1の機能をリン酸化することによってRP1の機能にブレーキをかけ、Mak(繊毛を短くする機能)とRP1(繊毛を長くする機能)のバランスを取ることで、繊毛の長さを調節していることも新たに解明しました(図5)。
本研究は、大阪バイオサイエンス研究所 発生生物学部門の大森 義裕 副部長を中心として、大阪大学超高圧電子顕微鏡センターの梶村 直子 研究員、名古屋大学 大学院医学系研究科の近藤 峰生 准教授の研究グループと共同で行われました。
<今後の展開>
この研究で、神経細胞のアンテナである繊毛の長さ調節機構の破綻が網膜色素変性症の原因となりうることを世界で初めて発見しました。近年、繊毛の異常が、肥満、糖尿病、嚢胞(のうほう)腎症、精神機能異常など種々の疾患の原因となっている可能性が指摘されています。今後、網膜色素変性症においてRP1以外のさまざまな原因遺伝子産物と繊毛の長さ調節との関連を解析することで、繊毛機能が疾患とどのようにかかわっているかの研究を進めます。また、神経細胞における第2の入力装置とも呼ばれる繊毛からの情報をどのように神経細胞が情報処理しシナプスへと伝えているかも解明していきます。
<参考図>
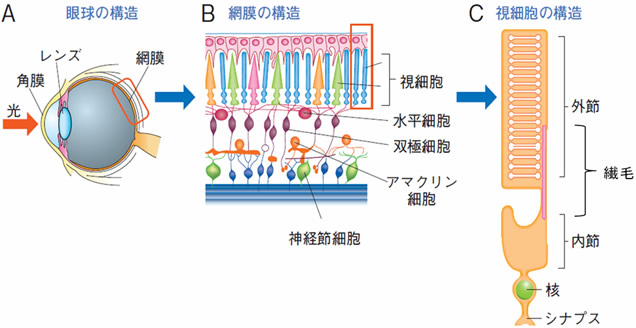
図1 網膜と視細胞の構造
- A: 網膜は目の後部に位置する膜状の組織で、角膜、レンズを通って入った光をキャッチする。
- B: 網膜では、視細胞と呼ばれる神経細胞がシート状に配列し、光センサーの役割を果たしている。
- C: 視細胞には、光をキャッチするアンテナの役割を果たす外節が存在するが、これは繊毛が発達した構造である。
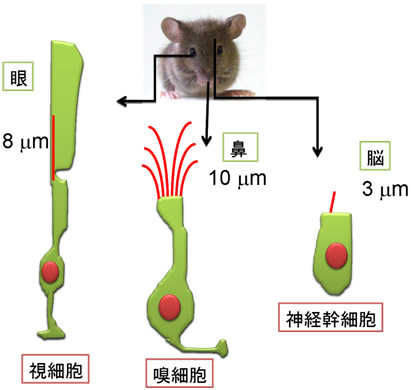
図2 さまざまな細胞の繊毛の長さ
細胞の種類により、繊毛の長さや本数は異なっている。例えば、視細胞は8μm程度の比較的長い繊毛を持つが、神経幹細胞は3μm程度の短い繊毛を持つ。
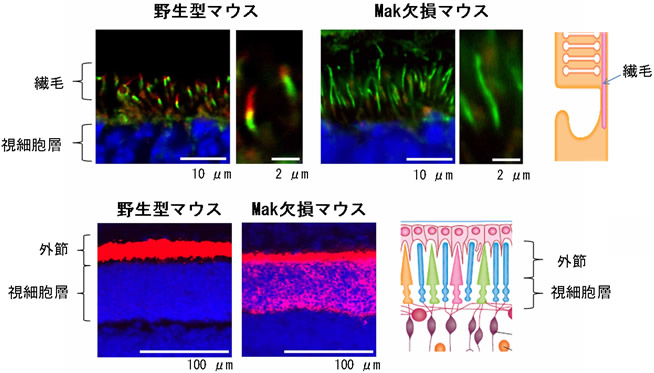
図3 野生型マウスとMak欠損マウスにおける繊毛の長さとロドプシン(視物質)の分布
マウス網膜に免疫染色を行った。
- 上段図: 繊毛(緑)が野生型に比べ、Mak欠損マウスでは伸びていることが分かる。
Mak(赤)は繊毛に局在する。
- 下段図: 野生型では外節にロドプシン(赤)の局在が見られるが、Mak欠損マウスでは視細胞層の細胞体にもロドプシンの蓄積が見られる。
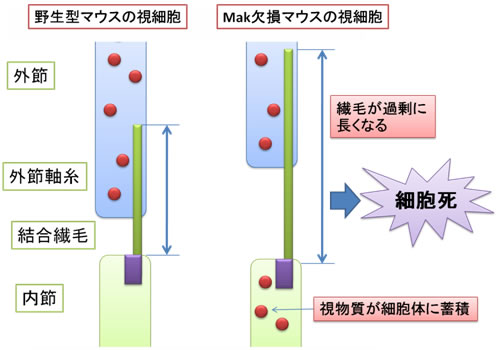
図4 野生型マウスとMak欠損マウスの視細胞の違い
Mak欠損マウスでは、視細胞の繊毛が過剰に長くなり、視物質が細胞体に蓄積することにより、視細胞が細胞死する。
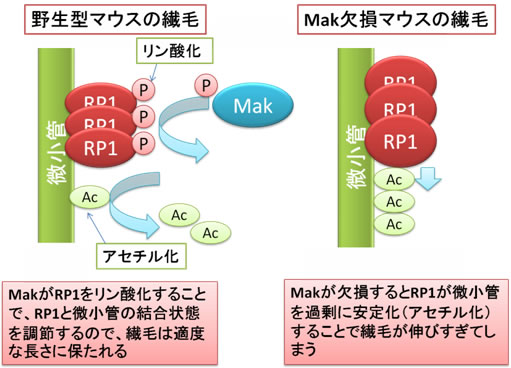
図5 Makによる繊毛の長さの制御機構
野生型マウスではMakはRP1をリン酸化することで、RP1と微小管の結合状態を調整し、繊毛は適度な長さに保たれる。Mak欠損マウスでは、RP1がリン酸化されず、微小管の過剰なアセチル化が引き起こされ、繊毛が伸びすぎてしまう。
<用語解説>
- 注1) 網膜色素変性症
- 網膜に異常が見られる遺伝性の病気で、人口3,000人から8,000人に1人の患者が存在する。夜盲、視野狭窄、視力低下が特徴的な症状である。
- 注2) 視細胞
- 網膜の最外層にあり、光を受け取り、電気信号に変換する細胞である。
- 注3) 繊毛
- 微小管の束からなる直径約0.25μmの小さな毛のような突出物で、さまざまな細胞の表面に形成される。光や匂い、尿の流れなどの外部情報をキャッチするためのアンテナとして機能することが知られており、目・鼻・耳・腎臓などで重要な役割をしている。また、精子やミドリムシの鞭毛も繊毛の一種である。繊毛の長さは細胞ごとに決まっているが、決まった長さに作られる仕組みはこれまでよく分かっていなかった。
- 注4) 視物質
- 光センサーの役割を担う光感受性分子。視細胞の外節に蓄えられ、光信号を化学シグナルに変換する一連の反応の発端となる。明暗を高感度に感知するロドプシン、特定の色を感知するコーンオプシンなどが知られる。
- 注5) スクリーニング
- ゲノムから目的の遺伝子または遺伝子群を探索すること。
<論文名>
“Negative regulation of ciliary length by ciliary kinase Mak is required for retinal photoreceptor survival”
(繊毛キナーゼMakによる繊毛の長さ調節は、網膜における視細胞の生存に必須である)
<お問い合わせ先>
<研究に関すること>
古川 貴久(フルカワ タカヒサ)
大阪バイオサイエンス研究所 発生生物学部門 研究部長
〒565-0874 大阪府吹田市古江台6-2-4
Tel:06-6872-4853 Fax:06-6872-3933
E-mail:
<JSTの事業に関すること>
河村 昌哉(カワムラ マサヤ)
科学技術振興機構 イノベーション推進本部 研究領域総合運営部
〒102-0075 東京都千代田区三番町5 三番町ビル
Tel:03-3512-3531 Fax:03-3222-2066
E-mail: