| タンパク質立体構造の柔軟性と高精度の動作機構 |
生命活動の場では、膨大な種類と数のタンパク質ナノマシンが会合離散を繰り返すことにより情報やエネルギーのやりとりを行い、複雑なネットワークを形成している。その信号処理、信号伝達機構は極めて高い信頼性と柔軟性を持つが、その基盤はタンパク質立体構造の設計原理にある。タンパク質はその柔軟性の高い立体構造のなかに、原子レベルの高精度動作機構、分子情報認識機構を持ち、特に複合体の場合は単なるナノマシンというより高度なシステムとして働き、複雑精緻な生命活動を支えている。
細菌べん毛は、個々のタンパク質分子から見れば巨大な複合体システムであり、その機械的・力学的動作を光学顕微鏡下で直接目にとらえることができるのが特徴である。直接観測されるべん毛の動作や柔軟性と、原子レベルの立体構造を結びつけることで、個々のタンパク質ナノマシンの動的構造物性や設計原理を明らかにすることが可能であると期待される。今回は一種類のタンパク質のみでらせん構造を形成するべん毛繊維の素繊維構造を原子レベルで解明し、その極微小かつ高精度のスイッチ機構を明らかにした。
| 細菌べん毛 |
細菌べん毛は細菌の運動器官であり、回転モータ、軸受、ユニバーサルジョイント、らせん型プロペラなど人工の推進機関と同等の機能を持つ部品から構成される複雑なタンパク質分子機械である。モータは細長いべん毛繊維を約20,000rpmという高速回転させ、それにより細菌は1秒間に体長の20倍ほどの距離を移動することができる。大腸菌やサルモネラ菌では、数本のべん毛が束になり同期して回転し、強い推進力を発生する。
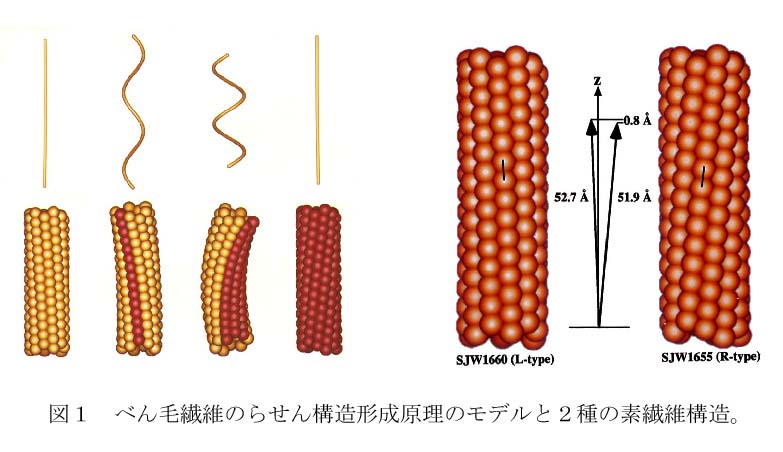
べん毛繊維は直径20nm、長さ10数ミクロンの細長い繊維で、11本の素繊維が円筒状に束になってできたチューブ構造である。素繊維はフラジェリンが一列に長く並んだもので、その並びの周期長がわずかに異なる2種類の構造間を切り替えることができ、それらが1本のべん毛に混在することで、べん毛は緩やかにねじれてらせんとなる。また、べん毛モータの急反転によりべん毛繊維にねじれ力が加わると、何本かの素繊維構造が切り替わって、らせんが左巻きから右巻きに変換する。まっすぐ泳ぐとき束になっていた数本のべん毛は、これによって直ちにほぐれ、推進力のアンバランスを生じて菌体は運動方向を変える。X線繊維回折法による精密測定から、2種類の素繊維構造がそれぞれ52.7Åと51.9Åという周期長を持ち、このわずか0.8Åという周期差が実際にべん毛らせんの構造設計に使われていることがわかっていた。すなわち、素繊維は0.1Å精度の力学的長さスイッチ機構を持つ。(図1)
べん毛繊維については、極低温電子顕微鏡やX線繊維回折法により10Å程度の分解能ですでに構造が得られており、サブユニットの外形、2次構造(アルファへリックスやベータ構造)の分布、配列様式などは明らかになっていたが、素繊維の高精度スイッチ機構を解明するには、原子レベルの詳細な構造決定が必要であった。そこで、サブユニットであるフラジェリンの結晶化によるX線結晶構造解析を試みた。
| 構造解析の方法 |
原子レベルでタンパク質の構造を決めるためには、分子が極めてきれいに並んだ結晶が必要である。フラジェリンは強い繊維重合能力を持ち、どのような結晶化でも必ず繊維を形成するため、結晶化ができなかった。そこで、サルモネラ菌のフラジェリン(分子量約51,500)を構成する494残基のアミノ酸のうち、繊維形成と構造安定化に重要な領域(N末端53残基、C末端44残基)を切り取り、繊維構造形成能力を失った分子量約41,000のフラグメント(F41と略記)を用いることにより初めて結晶を得ることに成功した。得られた結晶は厚さ数ミクロンの薄い板状であったため、データ収集は困難を極めたが、凍結結晶による低温結晶解析法を工夫したことと、第3世代放射光注1SPring-8の理研ビームライン(BL45XU)の強力なX線とトリクロメータ光学系による多波長異常分散(MAD)法を適用することにより、2.0Å分解能の回折データを得ることができた。それを解析して得られた電子密度のマップとアミノ酸配列のデータとを合わせて、F41分子の立体構造を決定した。
| 結晶中の素繊維構造と素繊維の力学的スイッチ機構 |
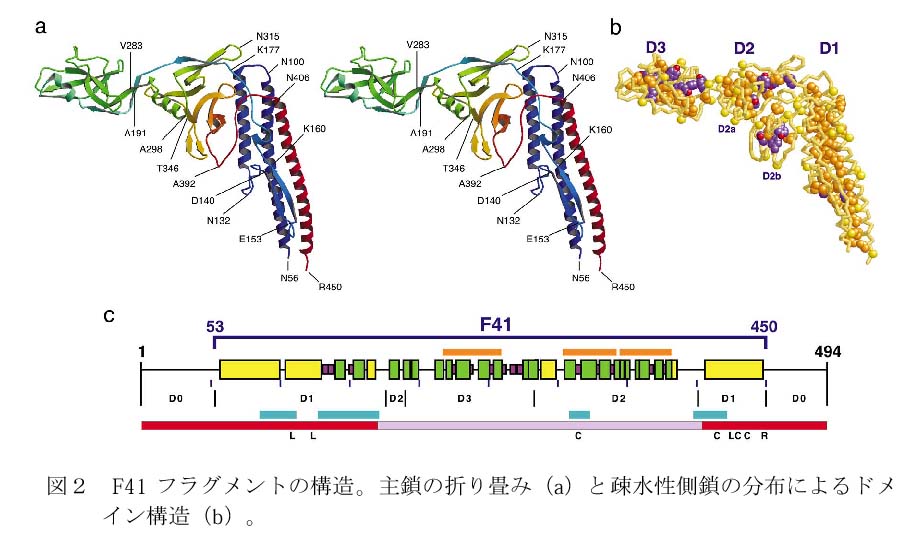
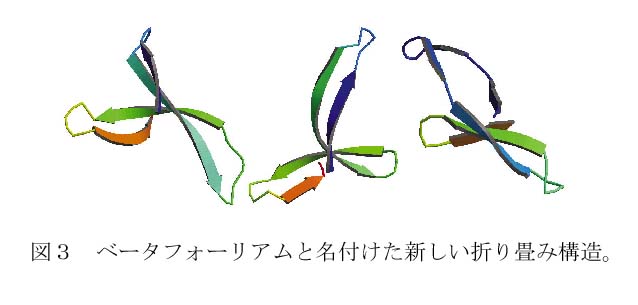
F41の分子構造は、図2に示すように3つのドメイン(領域)D1、D2、D3からなり、D1は3本の長いアルファヘリックスと1本のベータヘアピン注2で構成され、D2とD3はほとんどベータ構造でできている。D2とD3には、これまでに解明された数多くのタンパク質分子構造の中には見られないユニークな折り畳み構造(図3)も見られ、デカルトの葉線(フォーリアム)と呼ばれる3次曲線にちなんで、ベータフォーリアムと名付けた。
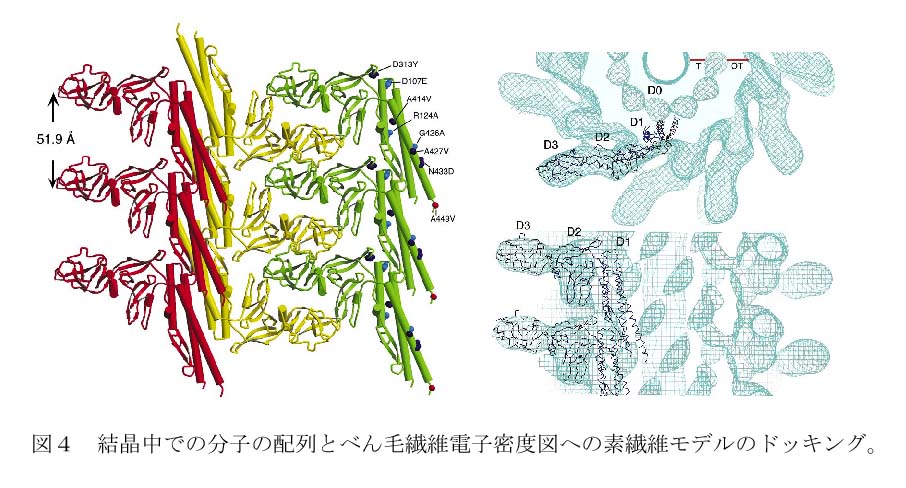
結晶構造のa軸に沿ったフラジェリンの繰り返し周期は51.9Åで、素繊維の繰り返し周期の1つと同じであった(図4左)。そこで、a軸に沿った分子の配列を取り出し、べん毛繊維の低分解能電子密度図に合わせてみると、図4右に示すように見事に一致した。つまり、べん毛繊維を形成する際には平行に束ねられてチューブを形成する素繊維が、結晶の中では反平行に並んでシートを形成し、このシートが積み重なって結晶となっていた訳である。
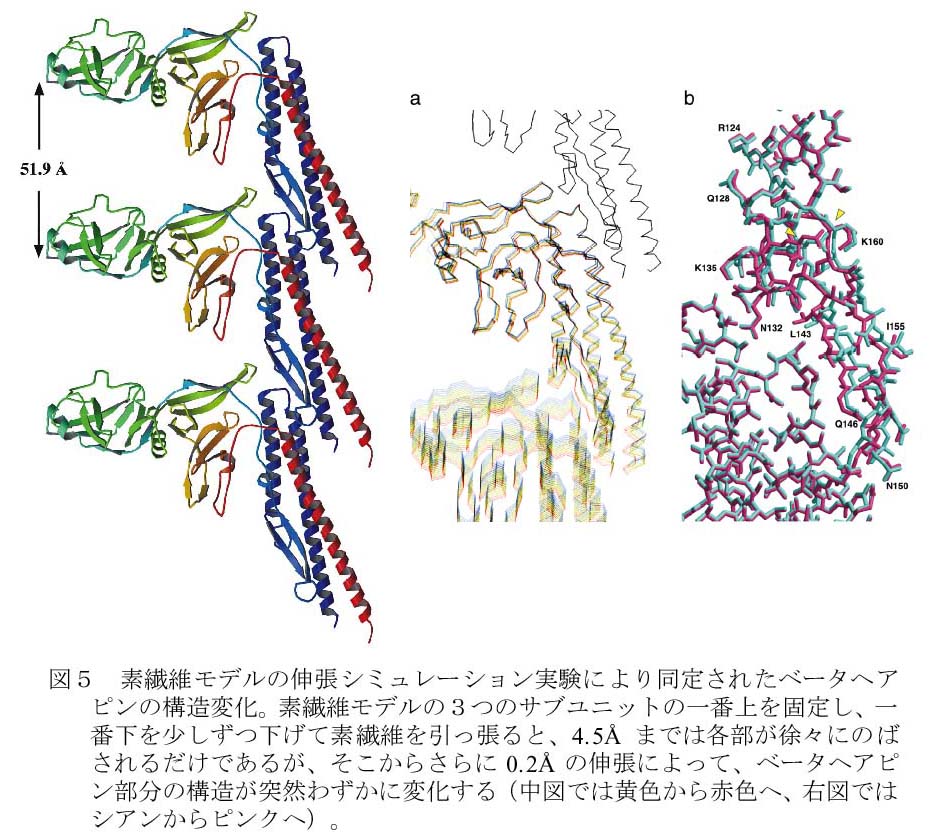
素繊維の高精度の力学的スイッチ機構を調べるため、決定された分子構造を用いた素繊維モデルに張力をかけるというシミュレーション実験を行った。具体的には、フラジェリン分子3個からなる素繊維を考え、一番上のフラジェリンを固定し、一番下のフラジェリンを0.1Åずつ下方に変位させながら中央の分子の構造変化を観察した。最初しばらくは徐々に構造が伸展しただけであったが、ドメインD1の分子間接触部位にあるのベータヘアピンが、あるところでわずかではあるが急激な構造変化を見せた(図5aとb)。このことから、異なる周期構造間を切り変わる素繊維の機械的・力学的スイッチが、このベータヘアピンの斜め下方向へのずれという、フラジェリン内部の微小な構造変化であることが明らかになった。
| 研究の意義と今後の展望 |
今回明らかにしたべん毛素繊維の0.1Å精度の力学的スイッチ機構は、タンパク質分子の立体構造がその成り立ちゆえに普遍的に持つ柔軟性と精密性との両立を見事に示したものである。生命活動を支える生物特有の柔らかく且つ高精度のしくみは、タンパク質が本来有する、こうしたナノマシンとしての高いポテンシャルに基づくものであることを示す。21世紀に向けてその基礎研究の発展と応用展開に大きな期待がかかるナノテクノロジーは、個々の原子を組み合わせ積み上げて、ナノスケールの機能素子や複雑な機能を持つナノマシンの構築を目指す研究分野であるが、生物の中にはそのお手本となるものが豊富にある。ただ、その高いポテンシャルを、我々はまだ十分に理解しているとは言い難い。さまざまな機能を持つタンパク質ナノマシンの構造と動作機構を明らかにし、動作原理についての理解をさらに深めることにより、ナノマシン設計原理に重要な指針が得られるであろう。
| 注1 | 第三世代放射光 |
| 兵庫県西播磨に建設されたSPring-8を始めとする最新の高輝度X線源のこと。高速で走る電子の軌道を曲げるときにエネルギーの一部が電磁波(光やX線)として放出される。これを放射光と呼び、電子の数が多いほど強い光が、電子の速度が大きいほど高いエネルギーの電磁波(X線)が放出される。SPring-8は世界最先端、最高性能の施設で、可視光からX線までの広い波長範囲で強力な光を出すことができる。バイオや材料科学を始め、広い分野の分析、研究あるいはプロセス開発に応用され、次々と画期的な成果をあげている。またこの施設によって初めて可能となった新しい手法も生まれつつある。 | |
| 注2 | アルファへリックスとベータヘアピン |
| アミノ酸が一列に結合したペプチド鎖がタンパク質の立体構造を作るための典型的な構成要素で、2次構造と呼ばれるものである。折れ畳まったペプチド鎖間で水素結合を形成し、安定かつ規則的な構造をなしているもの。らせん構造を作っているものをアルファへリックス、直線的な構造をベータストランドと呼ぶ。ベータストランドの折り返し構造がベータヘアピンである。 |
This page updated on March 15, 2001
Copyright©2001 Japan Science and TechnologyCorporation.
www-pr@jst.go.jp
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}