- JST トップ
- /
- 戦略的創造研究推進事業
- /
 ERATO
ERATO- /
- 研究領域の紹介/
- 終了領域/
- 長谷部分化全能性進化プロジェクト
長谷部分化全能性進化プロジェクト

研究総括 長谷部 光泰
(自然科学研究機構 基礎生物学研究所 教授)
研究期間:2005年~2011年
動物、植物ともにひとたび分化した細胞を幹細胞にプログラムしなおす(リプログラム)ことができます。植物は高いリプログラミング能力を持ち、簡単な処理だけで幹細胞を作れることが知られていました。しかし、その分子機構はほとんど明らかになっていませんでした。ヒメツリガネゴケはこれまでのモデル植物とは違い極めて高い分化全能性を持ち、陸上植物で最も容易に遺伝子ターゲティング法が行えます。我々はヒメツリガネゴケゲノム解読を出発点とし、分化した葉細胞を幹細胞に容易に変えることができる幹細胞誘導因子STEMIN遺伝子を発見しました。また、哺乳類の多能性幹細胞誘導を引き起こす因子がヒメツリガネゴケの幹細胞化も誘導するが、その分子機構は哺乳類と異なることもわかりました。これ以外にも多くの遺伝子がリプログラミングに必要であり、それらの相互作用を明らかにし、リプログラミング過程における遺伝子ネットワークの概要を明らかにしました。さらに、シダ植物ゲノム解読を通して、これらの因子が陸上植物全体の中でどのように進化したかを推定し、本プロジェクトの成果を将来的に品種改良などに利用していくことが可能であることがわかりました。
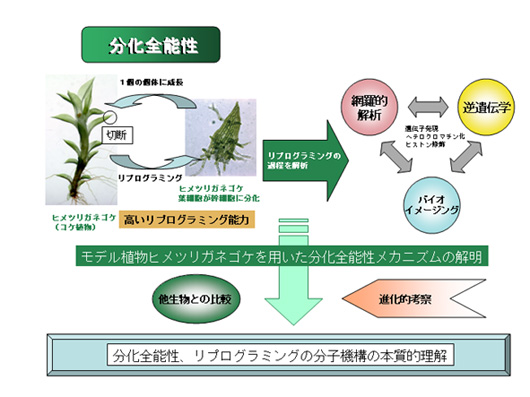
研究成果
インフォマティクス進化グループ
大量情報処理技術を用いて多能性幹細胞化過程のゲノム解析の基盤を整備するとともに、情報科学的に多能性幹細胞化のネットワークを解析し、多能性幹細胞化機構がどのように進化して来たかを解析した。本グループはヒメツリガネゴケゲノムプロジェクト、イヌカタヒバゲノムプロジェクトと連携しつつ両ゲノムのアノテーション、EST、cDNAの貼り付けによる遺伝子構造の同定、他の生物の遺伝子との関連づけ(オーソログ探索)をいち早く行い、両ゲノムと他生物ゲノムの比較解析を行った。とくに、全ゲノムのアセンブリー・アノテーションに先立ち、全ゲノムショットガンの生データから、他グループと共同で細胞周期制御、エピジェネティック遺伝子発現制御、光信号伝達、発生関連遺伝子について相同性に基づいて探索・同定した。また、上記の因子についてそれぞれ、遺伝子系統解析を行い、オーソログ・パラログ関係を推定した。
オミクスグループによるクロマチン修飾、遺伝子発現の動態を材料として、ネットワーク解析を行い、リプログラミング過程における遺伝子ネットワークを解析した。さらに、ネットワークの要素、相互作用を他の生物での知見と比較し、共通の祖先からリプログラミング機構がどのように進化してきたかを解析した。
- Rensing et al. (2008) The Physcomitrella genome reveals evolutionary insights into the conquest of land by plants. Science 319: 64.
- Banks, Nishiyama, Hasebe et al. (2011) The compact Selaginella genome identifies changes in gene content associated with the evolution of vascular plants. Science 332: 960.
バイオイメージンググループ
バイオイメージング技術を駆使し、多能性幹細胞化のアッセイ系、ならびに動態観察系を確立した。そして、生理学的・細胞生物学的視点から多能性幹細胞化に必要な因子の検討を行った。まず、効率的にアッセイを行うため、ライブイメージングにより多能性幹細胞化全過程を多検体同時観察できるシステムを構築した。次に、光、切断刺激、植物ホルモンなどの外部シグナルのうち、どのようなシグナルが多能性幹細胞化を制御するかを明らかにした。そして、これらの因子が幹細胞形成遺伝子ネットワークの中でどのような役割を果たすかを解明した。単独で幹細胞化を誘導できるSTEMINを始め、リプログラミングに重要な遺伝子を複数発見した。さらに、バイオイメージング技術によって多能性幹細胞化における細胞周期変動、ヒストン修飾関連因子の動態観察を行い、多能性幹細胞化における細胞周期再開とクロマチン修飾の役割を推定するべく研究を進めた。
逆遺伝学グループ
(1)逆遺伝学的実験方法の開発、(2)化学物質に対する表現型応答から原因遺伝子系を探索するケミカルジェネティクス法の確立、(3)それを用いた多能性幹細胞化に関わる遺伝子の単離と機能解析、そして、(4)多能性幹細胞化の鍵現象である細胞周期再開を引き起こす因子を逆遺伝学的手法により解明した。
まず、多能性幹細胞化に関わる遺伝子の機能解析を行うため、逆遺伝学的手法を開発した。具体的には、熱ショックプロモーター又はエストロジェン受容体を用いた条件的遺伝子発現誘導系、Cre-loxPシステムを用いた条件的遺伝子欠失系を構築した。さらに、約12,000種類の化学物質を用いてケミカルジェネティクス法により多能性幹細胞化を阻害あるいは誘導する因子を探索した。そして、ケミカルジェネティクス法により得られた候補遺伝子同士、ならびに他グループから得られた因子との相互作用を解析した。
また、多能性幹細胞化には細胞周期再開が伴うが、再開を引き起こす遺伝子ネットワークは動植物を通じて未解明な部分が多い。そこで、逆遺伝学的に、既知の細胞周期関連遺伝子をヒメツリガネゴケゲノム内から探索し、それらの遺伝子発現や相互作用を解析して、細胞周期再開の鍵となる細胞周期関連遺伝子を明らかにした。さらに、それらの遺伝子制御に関わる因子を探索し、細胞周期再開の分子機構を推定した。
- Ishikawa et al. (2011) Physcomitrella cyclin dependent kinase A links cell cycle reactivation to other cellular changes during reprogramming of leaf cells. Plant Cell 23: 2924.
オミクスグループ
ゲノムワイドな視点から、多能性幹細胞化過程における、クロマチン修飾とDNAメチル化変動、遺伝子発現制御の解析を行った。ヒメツリガネゴケにおいて、(1)超並列シーケンサーを用いたゲノムワイドなクロマチン修飾・DNAメチル化解析実験系、(2)マイクロアレイ、超並列シーケンサーを用いたmRNA・small RNA発現解析実験系を確立した。そして、多能性幹細胞化時における、クロマチン修飾変動、DNAメチル化変動、mRNA・small RNA発現変動を比較検討し、多能性幹細胞化に関与する候補因子を推定した。
上記の実験から得られた候補因子から、(a)多能性幹細胞化においてクロマチン修飾、DNAメチル化変動を引き起こす因子(クロマチン・DNA修飾酵素など)を探索し、同定した。(b)転写因子を選び、機能獲得又は機能喪失させることにより細胞周期再開に影響する因子を網羅的に選抜し、直接ターゲットを特定した。このために、高効率遺伝子機能解析実験系(アグロバクテリア形質転換系)、ゲノムワイドに直接ターゲットを効率的に特定する実験系(転写因子ChIP-seq法)を確立した。(c)small RNAによる(b)の転写因子の制御様式を推定した。(d)(a)から(c)のデータを統合し、多能性幹細胞化時における遺伝子発現制御ネットワーク変動を推定した。
- Nishiyama et al. (2012) Digital gene expression profiling by 5′-end sequencing of cDNAs during reprogramming in the moss Physcomitrella patens. PLoS ONE 7: e36471.











